Неисправности кпп: Неисправности механической коробки передач: выявление и возможности устранения
Неисправности механической коробки передач: выявление и возможности устранения
24.09.2014 #КПП # Коробка передач
Неисправности механической коробки передач: выявление и возможности устраненияМеханическая коробка передач и трансмиссия в целом — одна из наиболее нагруженных систем автомобиля, работающая в сложных условиях, поэтому для нее очень высока вероятность поломок. О том, какие неисправности характерны для механической КПП, как их диагностировать и устранить, а также об эксплуатации и обслуживании трансмиссии читайте в данной статье.
Основные неисправности МКПП и их причины
Трансмиссия предназначена для изменения и передачи крутящего момента от двигателя на колеса, то есть — это одна из главных систем автомобиля, определяющая его динамические и скоростные характеристики. Ключевую роль в трансмиссии играет коробка переключения передач (КПП, или просто коробка передач, КП), при выходе из строя которой автомобиль теряет свои ходовые качества, или вовсе не может двигаться.
Ключевую роль в трансмиссии играет коробка переключения передач (КПП, или просто коробка передач, КП), при выходе из строя которой автомобиль теряет свои ходовые качества, или вовсе не может двигаться.
Как известно, на сегодняшний день в мире существует два основных типа трансмиссии — ручная (механическая) и автоматическая. Эти трансмиссии имеют разное устройство и принцип действия, поэтому для них характерны разные неисправности. Здесь мы поговорим только о неисправностях механической коробки передач, о проблемах автоматической коробки передач читайте в отдельной статье.
Для механической коробки переключения передач (МКПП) характерны следующие неисправности:
• Шум во время работы и при переключении передач;
• Невозможность включения какой-либо передачи или всех передач;
• Затрудненное включение передач;
• Самопроизвольное выключение передач;
• Утечка масла из коробки передач.
Эти неисправности могут возникать вследствие износа шестерен, валов, подшипников, муфт синхронизаторов, шлицевых соединений, из-за потери герметичности сальников, вследствие самопроизвольного отворачивания болтов и гаек, при поломках в механизме привода и т.
Износ и поломки, в свою очередь, вызываются целым рядом причин:
• Естественный износ или выработка ресурса деталей;
• Использование некачественного трансмиссионного масла, или масла, отличного от рекомендованного производителем;
• Применение во время ремонта неоригинальных запчастей низкого качества;
• Нерегулярное техническое обслуживание или его отсутствие;
• Неквалифицированное обслуживание трансмиссии;
• Опасные и сложные для коробки передач режимы работы при спортивном и просто агрессивном стиле вождения.
Каждая неисправность трансмиссии проявляется теми или иными внешними признаками, однако для точного выявления проблемы необходимо провести диагностику.
Самостоятельная диагностика механической коробки передач
Проблема самостоятельной диагностики МКПП заключается в том, что обойтись здесь без ее снятия и разбора невозможно. Дело в том, что большинство неисправностей «механики» так или иначе происходит из-за износа деталей, а выявить и заменить изношенную шестерню или другую деталь можно только после разбора всей коробки. И только проблемы с приводом переключения передач можно решить без снятия коробки — для этого достаточно провести визуальный осмотр частей привода (рычагов, тяг, тросов, шарниров и т.д.), попробовать их работу и при необходимости заменить.
И только проблемы с приводом переключения передач можно решить без снятия коробки — для этого достаточно провести визуальный осмотр частей привода (рычагов, тяг, тросов, шарниров и т.д.), попробовать их работу и при необходимости заменить.
Порядок снятия механической коробки передач примерно одинаков для всех автомобилей:
• Поддомкратить или подвесить автомобиль (его переднюю часть) и коробку передач — это застрахует от падения и повреждения коробки после ее отсоединения от двигателя;
• Слить масло из коробки;
• На заднеприводном автомобиле — отсоединить карданный вал;
• На переднеприводном автомобиле — снять колеса, отсоединить ШРУСы и некоторые детали подвески;
• Открутить болты, удерживающие КПП на двигателе;
• Демонтировать крепления подвесных опорных подушек;
• Снять коробку передач;
• Разобрать коробку передач и провести диагностику.

При разборке КПП проверяется состояние шестерен, муфт синхронизаторов, шлицевых соединений валов, подшипников и других деталей, целостность всех уплотнителей и т.д. При выявлении неисправной детали производится ее замена, после чего коробка устанавливается на двигатель и заполняется трансмиссионным маслом.
Далеко не каждый из нас может или имеет желание самостоятельно снять, разобрать и проверить коробку передач, поэтому при неуверенности в своих силах лучше обратиться в сервис. И, конечно, в сервис обязательно нужно обращаться владельцам новых автомобилей, у которых еще не истек срок гарантии.
Неисправности механической коробки передач и их устранение
Для всех неисправностей коробки передач характерны те или иные внешние признаки — именно по этим проявлениям можно судить о характере поломки. Однако многие неисправности проявляются одинаково, поэтому в представленной здесь таблице все поломки МКПП сгруппированы по их внешним признакам.
| Неисправность МКПП | Причина неисправности |
|---|---|
| Коробка шумит при работающем двигателе и нейтральном положении рычага переключения передач | Падение уровня масла в КПП |
| Загрязнение или попадание воды в трансмиссионное масло | |
| Повреждение подшипника ведущей шестерни | |
| Износ или разрушение подшипника ведущего вала | |
| Шум при работающем двигателе на одной из передач | Поломка или износ синхронизатора |
| Разрушение, износ или поломка зубьев шестерни данной передачи | |
| Шум при работающем двигателе при включении всех передач | Те же причины, вызывающие шум на нейтральной передаче |
| Износ или разрушение подшипника выходного вала или подшипника ведомой шестерни | |
| Картер сцепления и коленчатый вал двигателя не соосны | |
| Шум в момент переключения передачи | Не до конца выключенное сцепление, неисправности сцепления |
| Износ или поломка муфт синхронизаторов | |
| Поломка блокирующего устройства или его деформация | |
| Самопроизвольное отворачивание болтов крепления коробки к двигателю | |
| Вибрации при работающем двигателе | |
| Самопроизвольное отворачивание болтов крепления коробки передач к двигателю или двигателя к кузову, разрушение опор крепления КПП и двигателя | |
| Утечка масла из коробки | Износ или повреждение масляных уплотнительных колец, сальников, пробок |
| Самопроизвольное отворачивание болтов крепления КПП к двигателю | |
| Чрезмерное количество масла в коробке | |
| Поломка или загрязнение сапуна | |
| Нарушение геометрии привалочных поверхностей КПП и картера сцепления (появление вмятин, забоев или трещин в результате ударов) | |
| Затрудненное или невозможное переключение передач | Не до конца выключено сцепление |
| Поломка или износ муфт синхронизаторов, появление на их внутренних сторонах задиров или заусенцев | |
| Просадка или поломка пружин синхронизаторов | |
| Заклинивание шестерен | |
| Нарушение в работе привода переключения передач (разрушение рычагов или тяг, поломка шарниров, заклинивание элементов и другие неисправности) | |
| Поломка блокирующего устройства | |
| Самопроизвольное отворачивание стопорных болтов на вилках | |
| Низкий уровень масла | |
| Загрязненное масло, попадание воды в масло | |
| Залито трансмиссионное масло, не соответствующее рекомендациям производителя | |
| Самопроизвольное выключение одной или нескольких передач | Износ или повреждение синхронизаторов |
| Износ или поломка шестерен | |
| Износ желобков на штоках вилок, шариков или пружин | |
| Самопроизвольное отворачивание болтов крепления КПП к двигателю или двигателя к кузову | |
| Износ или разрушение подшипников промежуточного или ведомого вала | |
| Потеря эластичности уплотнения рычага переключения передач | |
| Поломки в приводе переключения передач | |
| Отворачивание фиксатора подшипника на ведущей оси, поломка фиксатора этого же подшипника | |
| Износ шлицев на муфтах синхронизаторов | |
| Разрушение или износ вилки или штока переключения | |
| Просадка или повреждение пружин фиксаторов |
В данной таблице не указаны способы устранения неисправностей, так как они, в большинстве своем, требуют высокой квалификации и навыков ремонта автомобилей. Лишь некоторые операции может провести даже неподготовленный автовладелец — это замена и заливка масла, а также замена деталей привода включения передач. При обнаружении других неисправностей МКПП имеет смысл обратиться в автосервис.
Лишь некоторые операции может провести даже неподготовленный автовладелец — это замена и заливка масла, а также замена деталей привода включения передач. При обнаружении других неисправностей МКПП имеет смысл обратиться в автосервис.
Рекомендации по эксплуатации и обслуживанию МКПП
Долговечность и качество работы трансмиссии во многом зависит от того, как эксплуатируется коробка передач и сцепление. Продлить срок службы КПП, а значит — повысить комфорт, управляемость и безопасность автомобиля, несложно, для этого достаточно соблюдать несколько простых рекомендаций.
Главное в эксплуатации МКПП — плавное переключение передач с полным выжимом педали сцепления. Ни в коем случае не допускается переключение передач с неполным выключением сцепления, так как это приводит к ускоренному износу шестерен и повышает вероятность их поломки. Также очень важно двигаться на той передаче, которая наилучшим образом соответствует текущей скорости, а по возможности переключаться на прямую (обычно четвертую) передачу — в этом случае в передаче момента занято минимальное количество шестерен и два вала (в обход промежуточного), поэтому уменьшается износ деталей КПП.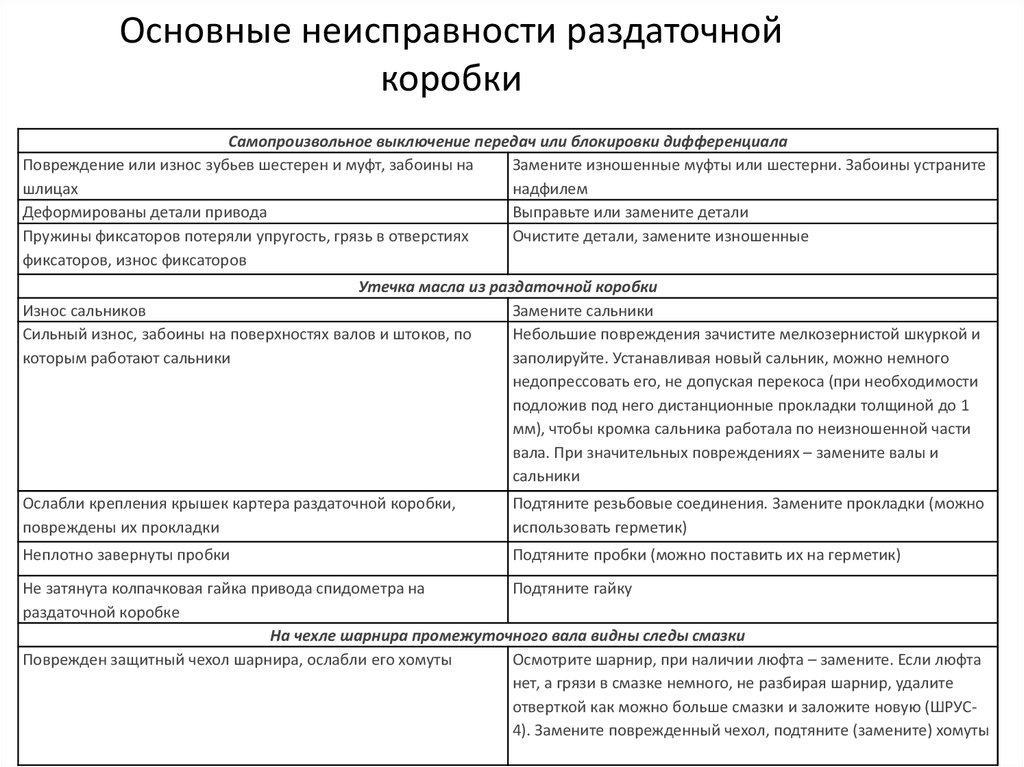
При переключении на пониженную передачу следует снижать скорость автомобиля, что также необходимо для снижения нагрузок на детали КПП и самого мотора. Ни в коем случае нельзя включать заднюю передачу даже при незначительной скорости движения автомобиля вперед — это обязательно приведет к повреждению КПП. И, наконец, если первая скорость не включилась сразу, то нужно сначала выжать сцепление (можно дважды), и лишь потом снова включить первую передачу. Далее переключение передач нужно выполнять плавно и с обязательной паузой между переключениями.
Механическая коробка передач имеет и особенности эксплуатации, о которых многие люди даже не догадываются. Например, в зимнее время года, особенно при значительных отрицательных температурах, не рекомендуется ставить автомобиль на стоянку с включенной передачей — в этом случае велик риск смерзания кольца синхронизатора, и при последующем пуске двигателя даже после выключения передачи автомобиль может тронуться. Также это чревато некоторыми неисправностями.
Большое значение для надежной работы коробки передач имеет и ее обслуживание. Во-первых, периодически после длительных и активных поездок нужно осматривать коробку передач на предмет появления потеков масла. Обычно потеки могут появляться в области заливной и сливной пробок, по периметру стыковки коробки к двигателю, между картером и крышкой КПП, а также (в случае зеднеприводного автомобиля) на фланце выходящего их коробка карданного вала.
Во-вторых, следует проверять уровень масла и при необходимости доливать его. Обычно проверка уровня масла проводится каждые 10-15 тысяч км пробега, а полная замена масла производится каждые 50-75 тысяч км пробега. Межсервисный интервал зависит от возраста автомобиля, в большинстве новых автомобилей в первые два-три года (а то и первые пять лет) масло вовсе не доливается, но далее проверку и долив масла нужно производить не реже, чем через каждые 20 тысяч км пробега (то есть, не реже одного раза в год).
Очень важно использовать рекомендованное производителем КПП трансмиссионное масло, и нельзя смешивать масла с разными характеристиками. В противном случае возможен интенсивный износ деталей коробки и ее выход из строя.
В противном случае возможен интенсивный износ деталей коробки и ее выход из строя.
При правильной эксплуатации и грамотном обслуживании механическая трансмиссия будет работать долго и надежно, не доставляя проблем водителю.
Другие статьи
#Стойка стабилизатора Nissan
Стойка стабилизатора Nissan: основа поперечной устойчивости «японцев»22.06.2022 | Статьи о запасных частях
Ходовая часть многих японских автомобилей Nissan оснащается стабилизатором поперечной устойчивости раздельного типа, соединенным с деталями подвески двумя отдельными стойками (тягами). Все о стойках стабилизатора Nissan, их типах и конструкции, а также о подборе и ремонте — читайте в данной статье.
#Ремень приводной клиновой
Ремень приводной клиновой: надежный привод агрегатов и оборудования
15.06.2022 | Статьи о запасных частях
Для привода агрегатов двигателя и в трансмиссиях различного оборудования широко применяются передачи на основе резиновых клиновых ремней. Все о приводных клиновых ремнях, их существующих типах, особенностях конструкции и характеристиках, а также о правильном выборе и замене ремней — читайте в статье.
Все о приводных клиновых ремнях, их существующих типах, особенностях конструкции и характеристиках, а также о правильном выборе и замене ремней — читайте в статье.
Барабан тормозной ГАЗ: управляемость и безопасность горьковских автомобилей
08.06.2022 | Статьи о запасных частях
Тормозные системы большинства ранних и актуальных моделей автомобилей ГАЗ оснащаются колесными механизмами барабанного типа. Все о тормозных барабанах ГАЗ, их существующих типах, конструктивных особенностях и характеристиках, а также о выборе, замене и обслуживании данных деталей — читайте в статье.
#Палец поршневой
Палец поршневой: прочная связь поршня и шатуна
02.02.2022 | Статьи о запасных частях
В любом поршневом двигателе внутреннего сгорания присутствует деталь, соединяющая поршень с верхней головкой шатуна — поршневой палец. Все о поршневых пальцах, их конструктивных особенностях и способах установки, а также о верном подборе и замене пальцев различных типов подробно рассказано в статье.
Вернуться к списку статей
Неисправности КПП у Honda — устранение дефектов и ремонт коробки передач
ВНИМАНИЕ! Изменения в графике работы магазина и сервиса в СПб: воскресенье — ВЫХОДНОЙ!
Коробка переключения передач
Коробка передач предназначена для изменения сил тяги на ведущих колесах и скоростей движения автомобиля путем увеличения или уменьшения передаточного числа.
В зависимости от характера изменения передаточного числа различают коробки:
- ступенчатые
- бесступенчатые
- комбинированные
По характеру связи между ведущим и ведомым валами коробки передач делят на:
- Механические
- Гидравлические
- Электрические
- Комбинированные
По способу управления на:
- Автоматические
- Полуавтоматические
- Неавтоматические
Коробки передач — наиболее устойчивая часть автомобиля. Поломки КПП встречаются гораздо реже, чем других частей автомобиля. Главный фактор поддержания коробки передач в исправном состоянии — хорошее состояние масла в коробке.
Поломки КПП встречаются гораздо реже, чем других частей автомобиля. Главный фактор поддержания коробки передач в исправном состоянии — хорошее состояние масла в коробке.
Все коробки передач очень близки по своим показателям. Переднеприводные машины с цилиндрическими главными передачами, расположенными прямо в картере коробки, позволяют использовать в трансмиссии автомобиля то же масло, что используется и в двигателе.
Коробки передач заднеприводных автомобилей заправляются специальным трансмиссионным маслом.
Механические коробки передач более просты в конструкции, не требуюют специальных гидравлических жидкостей для работы гидротрансформатора, менее склонны к отказам.
Автоматические коробки выигрывают удобством эксплуатации, их ремонт хорошо освоен на СТО. Автоматическая коробка передач на подержанной иномарке — это необходимый элемент риска. Такая трансмиссия накладывает на водителя повышенные обязательства по соблюдению правил эксплуатации.
Основные неисправности в коробке передач и их устранение
| Причина поломки | Устранение или предотвращение |
| Шум в коробке передач | |
| Износ подшипников, зубьев шестерен и синхронизаторов или их поломка | Заменить изношенные детали |
| Недостаточный уровень масла в коробке передач | Долить масло. Проверить и при необходимости устранить причины утечки масла |
| Некачественное масло в коробке передач | Заменить масло |
| Осевое перемещение валов | При необходимости заменить детали, фиксирующие подшипники или сами подшипники |
| Износ втулок оси шлицевого вала заднего хода | Заменить втулки шлицевого вала |
| Затруднённое переключение передач | |
| Неполное выключение сцепления | Проверка и ремонт сцепления |
| Заедание поверхности сферического шарнира | Снять рычаг и зачистить сопрягающиеся поверхности сферического шарнира |
| Деформация рычага переключения передач | Снять рычаг, устранить деформацию или заменить рычаг |
| Тугое движение штоков вилок (заусеницы, загрязнение, заклинивание блокировочных сухарей) | Разобрать, выявить причину, при необходимости отремонтировать или заменить изношенные детали |
| Неисправность синхронизаторов | Заменить изношенные детали или синхронизатор в сборе |
| Картер заправлен маслом несоответствующей марки | Слить масло, промыть коробку передач и заправить маслом или маслом-заменителем, рекомендуемым производителем |
| Деформация вилок привода переключателя | Выправить вилки, при необходимости заменить |
| Ослабление затяжки или отвертывание винтов головок механизма переключения | Завернуть и закрепить винты |
| Разбиты отверстия под штифты в горловине механизма переключения | Заменить крышку механизма переключения или отремонтировать, расточив отверстия и запрессовав ступенчатые штифты |
| Самопроизвольное выключение или нечёткое включение передач | |
| Неправильное включение передач | При выжатой педали сцепления рычаг переключения перемещать до упора |
| Износ шариков или потеря упругости пружин фиксаторов штоков переключения передач | Снять крышку фиксаторов и осмотреть детали; при необходимости заменить |
| Износ или неправильное положение блокировочных сухарей штоков переключения передач | Разобрать и заменить изношенные детали, следя за правильностью сборки |
| Износ блокирующих колец синхронизаторов | Заменить изношенные кольца синхронизатора |
| Поломка пружин синхронизаторов | Заменить пружины |
| Износ зубьев муфты синхронизатора или зубчатого синхронизатора шестерни | Заменить муфту или шестерни |
| Ослабление затяжки гаек крепления коробки передач к картеру сцепления или гаек крепления удлинителя к картеру коробки передач | Затянуть гайки |
| Износ вкладышей управления переключением или износ резиновых деталей в рычагах переключения передач | Заменить изношенные детали |
Замена трансмиссионного масла
Как и моторное масло трансмиссионное масло подвержено процессу старения.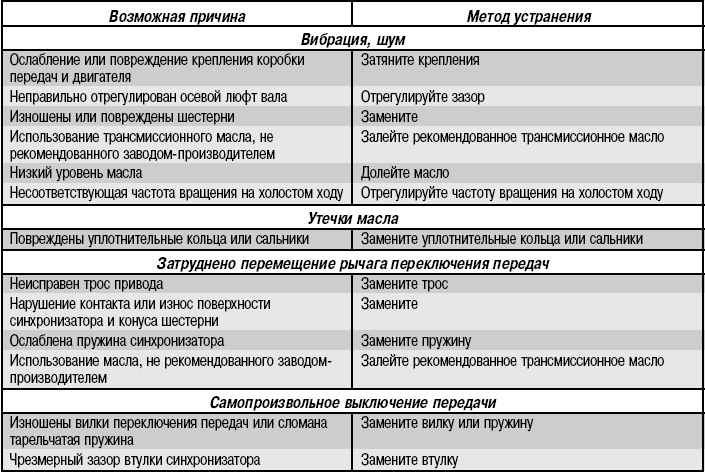 Оно теряет свои первоначальные свойства и перестает выполнять свои функции.
Оно теряет свои первоначальные свойства и перестает выполнять свои функции.
Поддержание трансмиссионного масла в хорошем состоянии значительно продлевает срок жизни КПП. Для этого необходимо выполнение как минимум 2 пунктов:
- Своевременная замена трансмиссионного масла
- Применение промывки масла перед его заменой
Промывка — важная часть процесса замены масла
При смене масла без применения промывки значительная часть загрязнений остается в КПП.
Промывка:
- Размягчает и удаляет нагар, продукты износа, углеродистые отложения
- Прочищает масляные каналы, улучшая циркуляцию масла
- Обеспечивает более полный слив старого масла
- Безопасна для резиновых уплотнителей, сальников
Контрольные точки клеточного цикла и анеуплоидия на пути к раку
In Vivo. 2018 январь-февраль; 32(1): 1–5.
Опубликовано в Интернете 15 января 2018 г. doi: 10. 21873/invivo.11197
21873/invivo.11197
Информация об авторе Примечания к статье Информация об авторских правах и лицензии Отказ от ответственности регуляторные белки для правильного клеточного размножения, включая циклиновые белки и циклинзависимые киназы, онкогены и гены-супрессоры опухолей, а также белки митотических контрольных точек. Мутации любого из этих регуляторных механизмов могут привести к репродукции клеток, несущих генетические мутации или аномальное число хромосом, что приводит к нестабильности генома. Хромосомная нестабильность, способствующая нестабильности генома, относится к аномалиям числа хромосом и приводит к анеуплоидии. Роль анеуплоидии в развитии раковых клеток часто оспаривается, поскольку противоречивые гипотезы и исследования не позволяют понять, является ли анеуплоидия причиной или следствием рака. Здесь мы представляем обзор важности регуляции контрольных точек клеточного цикла и хромосомной нестабильности в развитии рака и обсуждаем доказательства противоречивых аргументов в пользу роли анеуплоидии в развитии рака, что приводит нас к выводу, что дальнейшее изучение этой роли принесло бы пользу. наше понимание развития рака.
наше понимание развития рака.
Ключевые слова: Контрольные точки клеточного цикла, рак, регуляция, циклины, циклинзависимые киназы, анеуплоидия, обзор одинаковые дочерние клетки. Этот цикл и его регуляция необходимы для роста и размножения клеток и включают в себя множество регуляторных белков, таких как циклиновые белки и циклинзависимые киназы (CDK), онкогены и гены-супрессоры опухолей в интерфазе, а также белки контрольных точек митоза, которые обеспечивают стадии клеточный цикл продолжаться или тормозить этот процесс. Однако мутации и недостатки в регуляции на протяжении клеточного цикла могут привести к серьезным заболеваниям, таким как рак. В этом обзоре клеточный цикл в нормальных клетках сравнивается с клеточным циклом в раковых клетках с акцентом на задействованные регуляторные белки и роль дефицита этих белков в развитии рака.
Клеточный цикл состоит из интерфазы, состоящей из фаз G 1 , S и G 2 , и митотической (М) фазы (1). Во время интерфазы клетка готовится к делению посредством роста и репликации ДНК. Фаза G 1 представляет собой промежуток между окончанием цитокинеза предыдущего деления и началом S-фазы и представляет собой фазу, в которой клетка растет, готовясь к репликации ДНК, а также фазу, в которой принимается решение будет ли клетка снова делиться или войдет в G 0 , фаза покоя. Удаление факторов роста в начале G 1 отправит клетку в G 0 , но их удаление позже в G 1 , после контрольной точки рестрикции, позволит клетке продолжить S-фазу (2). Во время S-фазы происходит репликация ДНК, и каждая хромосома удваивается, превращаясь в две сестринские хроматиды. G2 отмечает промежуток между концом S-фазы и началом митоза. Здесь клетка синтезирует материалы, необходимые для митоза, такие как РНК и белки (3).
Во время интерфазы клетка готовится к делению посредством роста и репликации ДНК. Фаза G 1 представляет собой промежуток между окончанием цитокинеза предыдущего деления и началом S-фазы и представляет собой фазу, в которой клетка растет, готовясь к репликации ДНК, а также фазу, в которой принимается решение будет ли клетка снова делиться или войдет в G 0 , фаза покоя. Удаление факторов роста в начале G 1 отправит клетку в G 0 , но их удаление позже в G 1 , после контрольной точки рестрикции, позволит клетке продолжить S-фазу (2). Во время S-фазы происходит репликация ДНК, и каждая хромосома удваивается, превращаясь в две сестринские хроматиды. G2 отмечает промежуток между концом S-фазы и началом митоза. Здесь клетка синтезирует материалы, необходимые для митоза, такие как РНК и белки (3).
Фаза М начинается с митоза, который подразделяется на пять фаз, а именно профазу, прометафазу, метафазу, анафазу и телофазу, и заканчивается цитокинезом (1). Размножение через клеточный цикл приводит к генетически идентичным дочерним клеткам.
Размножение через клеточный цикл приводит к генетически идентичным дочерним клеткам.
На протяжении всех этих фаз клеточного цикла регуляция необходима для правильного производства дочерних клеток. Фундаментальным аспектом регуляции являются циклиновые белки и циклинзависимые киназы (CDK), которые при активации образуют комплексы и катализируют продвижение по клеточному циклу (2). В результате систематического синтеза и разрушения циклина на протяжении всего цикла CDK активируются только в определенные моменты клеточного цикла, что является ключевым фактором регуляции клеточного цикла (4). После активации циклинами CDK фосфорилируют специфические субстраты, которые управляют событиями клеточного цикла и клеточного деления.
Эти регуляторные роли комплексов циклин-CDK позволяют создавать «контрольные точки» во время клеточного цикла. Контрольные точки клеточного цикла отвечают за то, чтобы каждый предыдущий процесс был завершен до того, как ячейка перейдет к следующей фазе цикла. Активация контрольной точки, означающая обнаружение возможной ошибки, останавливает клеточный цикл в его текущей фазе за счет изменений уровней CDK и активации, предотвращая неправильное клеточное размножение. Регулирующие контрольно-пропускные пункты включают G 1 /S или контрольная точка рестрикции, G 2 /M или контрольная точка репликации ДНК и контрольная точка метафазы/анафазы или веретенообразного аппарата. На контрольную точку рестрикции в первую очередь влияют факторы роста, размер клеток, питание клеток и повреждение ДНК (2,5). Контрольная точка репликации ДНК в первую очередь зависит от неправильной репликации и повреждения ДНК, а контрольная точка метафазы/анафазы зависит от прикрепления хромосомы к митотическому веретену (1). Если какие-либо процессы будут обнаружены незавершенными или в этих контрольных точках будут очевидны повреждения, регуляторная активность циклин-CDK блокируется, что препятствует продвижению клетки по циклу до тех пор, пока эти проблемы не будут решены и клетка не будет готова к следующей фазе (1,6).
Активация контрольной точки, означающая обнаружение возможной ошибки, останавливает клеточный цикл в его текущей фазе за счет изменений уровней CDK и активации, предотвращая неправильное клеточное размножение. Регулирующие контрольно-пропускные пункты включают G 1 /S или контрольная точка рестрикции, G 2 /M или контрольная точка репликации ДНК и контрольная точка метафазы/анафазы или веретенообразного аппарата. На контрольную точку рестрикции в первую очередь влияют факторы роста, размер клеток, питание клеток и повреждение ДНК (2,5). Контрольная точка репликации ДНК в первую очередь зависит от неправильной репликации и повреждения ДНК, а контрольная точка метафазы/анафазы зависит от прикрепления хромосомы к митотическому веретену (1). Если какие-либо процессы будут обнаружены незавершенными или в этих контрольных точках будут очевидны повреждения, регуляторная активность циклин-CDK блокируется, что препятствует продвижению клетки по циклу до тех пор, пока эти проблемы не будут решены и клетка не будет готова к следующей фазе (1,6). .
.
В частности, три CDK участвуют в регуляции во время интерфазы (CDK2, CDK4 и CDK6), регулируя выход из субфазы и вход в нее (7). Обнаружение повреждения ДНК сигнализирует об ингибировании этих CDK, вызывая остановку клеточного цикла. Активация CDK4 и CDK6 влияет на раннее развитие G 1 , связываясь с циклином-D для фосфорилирования pRb, белка ретинобластомы, предотвращая его связывание и ингибируя фактор транскрипции E2F, который транскрибирует необходимые белки для G 9.0017 1 /S переход и продвигает клетку к следующим фазам цикла. pRb отвечает за инактивацию E2F во время таких фаз, как G 0 и M. Когда CDK4 и CDK6 ингибируются при обнаружении повреждения ДНК, они не инактивируют pRb, позволяя ему связываться и ингибировать E2F и предотвращая транскрипцию. необходимых белков, процесс, который возобновляется, когда контрольная точка считает, что клетка была должным образом подготовлена (7). CDK2 также участвует в инактивации pRb, а также играет роль в репарации и репликации ДНК, фосфорилируя субстраты, необходимые для репликации ДНК (8,9). ).
).
Онкогены и гены-супрессоры опухолей играют большую роль в регуляции клеточного цикла, особенно путей p53 и pRb, которые участвуют в контрольной точке рестрикции.
p53, белок-супрессор опухоли, реагирует на повреждение ДНК во время G 1 , такое как несоответствие и одноцепочечная ДНК, инициируя транскрипцию p21 , белка, взаимодействующего с CDK, который ингибирует активацию необходимых CDK G1 для фосфорилировать pRb и, следовательно, предотвращать прогрессирование синтеза ДНК (11). р53 также принимает активное участие в апоптозе в случаях непоправимого повреждения (12). Мутации в p53 чрезвычайно распространены при раке человека и приводят к отсутствию транскрипции p21 и, следовательно, дисфункции или отсутствию ареста G1 при наличии повреждения ДНК, что позволяет этому повреждению продолжаться в цикле без восстановления (13). Доказательства этой зависимости от p53 во время G 1 были продемонстрированы в исследованиях p53 -null трансгенных мышей и неспособности их клеток останавливаться во время G 1 при столкновении с повреждением ДНК (14). В результате мутация или повреждение становятся постоянными в геноме (2). Исследования мутаций в p53 , которые часто сопровождаются конформационными изменениями белка, которые позволяют обнаруживать эти мутации (11), у мышей было показано, что мутантный p53 приводит к восприимчивости к опухолям, таким как аденокарциномы легких, остеосаркомы и лимфомы (15).
В результате мутация или повреждение становятся постоянными в геноме (2). Исследования мутаций в p53 , которые часто сопровождаются конформационными изменениями белка, которые позволяют обнаруживать эти мутации (11), у мышей было показано, что мутантный p53 приводит к восприимчивости к опухолям, таким как аденокарциномы легких, остеосаркомы и лимфомы (15).
Связанный с опухолевым супрессором p53, мышиный онкоген двойной минуты-2 ( MDM2 ) также участвует в контрольной точке повреждения ДНК. Белки MDM2 устраняют контрольную точку, связываясь с мутантным p53 дикого типа и ингибируя его функцию посредством этих взаимодействий (16,17).
pRb, обсуждавшийся ранее, является еще одним супрессором опухоли и мишенью CDK (CDK2, CDK 4 и CDK6), участвующих в G1 и контрольной точке рестрикции (5,18). Дефектная функция pRb приводит к отсутствию связывания с E2F, который затем позволяет транскрибировать такие белки без регуляции и продвигать клетку по циклу (19).
Было показано, что циклин D1 амплифицируется при раке, таком как рак молочной железы (20). Циклин D1 влияет на функцию pRb посредством связывания с CDK 4 и CDK6, которые фосфорилируют pRb.
Циклин D1 влияет на функцию pRb посредством связывания с CDK 4 и CDK6, которые фосфорилируют pRb.
Другой источник развития рака и онкогенеза лежит в дефектной регуляции во время митоза. Анеуплоидия связана с наличием аномального числа хромосом в клетке и является чрезвычайно распространенной характеристикой опухолевых клеток с хромосомной нестабильностью (21, 22). Хромосомная нестабильность относится к повышенной частоте хромосомных аномалий, таких как делеции или дупликации, что приводит к неравному распределению ДНК в дочерних клетках. Высокие показатели хромосомной нестабильности могут привести к анеуплоидии, которая часто наблюдается при раке. Например, прирост целых хромосом, такой как прирост хромосомы 8, был обнаружен как распространенная ошибка в кариотипе при остром миелоидном лейкозе (23).
Одна из гипотез относительно того, почему это аномальное расхождение хромосом может привести к развитию опухоли, заключается в том, что если в клетке отсутствуют части хромосом или целые хромосомы, они также теряют регуляторные гены, включенные в эти части, такие как гены-супрессоры опухоли (24). Эти аномальные числа также могут приводить к аберрантной экспрессии генов и различным уровням геномной нестабильности (25).
Эти аномальные числа также могут приводить к аберрантной экспрессии генов и различным уровням геномной нестабильности (25).
Исследования показывают, что нормальная клетка, такая как клетка из клеточной линии RPE-1 или HCT116, неправильно сегрегирует хромосому один раз на каждые сто циклов деления (26). Считается, что эти неправильные сегрегации вызваны ошибками, окружающими контрольную точку веретена, такими как аномальные числа центросом и неправильное прикрепление кинетохор к микротрубочкам. Напр., меротелиальные прикрепления, при которых одна единственная кинетохора пары сестринских хроматид прикрепляется к микротрубочкам с обоих полюсов клетки, могут возникать из-за избыточного числа центросом (27). Ганем и др. предположили, что посредством генерации и визуализации клеток, отличающихся числом центросом, дополнительные хромосомы приводят к высокой частоте меротелиальных прикреплений и неправильной сегрегации, ведущих к анафазе (27). Особенно важно изучить механизмы, окружающие контрольную точку веретена, поскольку клеточный цикл зависит от этой контрольной точки, чтобы обеспечить правильное расхождение хромосом перед делением.
Перед началом анафазы и хромосомной сегрегации сестринские хроматиды удерживаются вместе с помощью белкового комплекса cohesin (28). Протеолиз слипчивости сестринских хроматид 1 (SCC1), субъединицы когезина, сепаразой, протеазой, запускает анафазу и хромосомную сегрегацию. Как только подтверждено правильное прикрепление хромосом к волокнам веретена деления, ингибитор сепаразы, называемый секурином, убиквитинируется комплексом, способствующим анафазе, или циклосомой APC/C. Без секурина, ингибирующего сепаразу, она активируется, происходит протеолиз SCC1 и начинается анафаза (28). Считается, что при обнаружении неправильного соединения хромосом с аппаратом веретена на кинетохорах эти кинетохоры посылают сигнал контрольной точки для остановки клеточного цикла, сигнализируя об ингибировании APC/C и, следовательно, подавляя сегрегацию (29).,30). Исследования, включающие лазерную абляцию одной неприкрепленной кинетохоры, показали, что клетка может продолжать митоз, когда происходит абляция, указывая на то, что неприкрепление является источником ингибирующего сигнала (29).
В то время как полный отказ в этой контрольной точке веретена ведет к гибели клеток (31), в опухолевых клетках были обнаружены нарушения, но не отказ контрольной точки, возникающие из-за дефектов, окружающих белки, участвующие в этом процессе. Эти белки после выявления неправильного прикрепления хромосом к аппарату веретена ингибируют прохождение стадий, окружающих АРС/С, тем самым предотвращая инициацию и продвижение сегрегации и анафазы. Способность APC/C к убиквитинированию зависит от связывания белка 20 цикла клеточного деления (CDC20), который рекрутирует субстраты для APC/C и активирует процесс (32). Другие белки, участвующие в функционировании контрольной точки веретена, включают митотический дефицитный белок 2 (MAD2) и почкование, не ингибируемое бензимидазол-родственным белком 1 (BUBR1). MAD2 связывается с CDC20, ингибируя белок и предотвращая убиквитинирование секурина с помощью APC/C (33). BUBR1 также ингибирует комплекс APC/C и CDC20 независимо от MAD2 (33). Изменения в экспрессии этих белков контрольных точек могут привести к неправильному расхождению хромосом из-за невозможности остановить их до того, как можно будет внести исправления, что приведет к анеуплоидии. Эти аберрантные числа хромосом часто наблюдаются при раке, о чем свидетельствует общность дефицита контрольных точек веретена в опухолевых клетках (22).
Эти аберрантные числа хромосом часто наблюдаются при раке, о чем свидетельствует общность дефицита контрольных точек веретена в опухолевых клетках (22).
Мутации в белках контрольных точек часто обнаруживаются в связи с колоректальным раком, в опухолевых клетках которого обычно обнаруживаются признаки анеуплоидии из-за хромосомной нестабильности (22,31,34). Кроме того, Li и Benezra сообщили, что линия клеток рака молочной железы со сниженным уровнем MAD2 (T47D) не останавливалась в контрольной точке веретена в ответ на ингибирующее воздействие на аппарат веретена (35). Мутации или дисрегуляция других белков контрольных точек, таких как почкование, не ингибируемое бензимидазольным белком 1 (BUB1) и BUBR1, также наблюдались при колоректальном раке и раке легкого, а также при лейкемии и лимфоме (36-38). Джемма и др. провел скрининг последовательностей ДНК клеточных линий рака легкого человека на наличие мутаций в гене BUB1 и обнаружил различные изменения этого гена в некоторых из этих клеточных линий (36). Ошима и др. обнаружили мутации или делеции в генах BUB1 и BUBR1 при Т-клеточном лейкозе/лимфоме у взрослых (37). Ситири и др. обнаружили нарушения в экспрессии BUB1 и BUBR1 при колоректальном раке и пришли к выводу, что аномальная экспрессия этих генов контрольных точек может приводить к анеуплоидии и метастазированию опухоли (38).
Ошима и др. обнаружили мутации или делеции в генах BUB1 и BUBR1 при Т-клеточном лейкозе/лимфоме у взрослых (37). Ситири и др. обнаружили нарушения в экспрессии BUB1 и BUBR1 при колоректальном раке и пришли к выводу, что аномальная экспрессия этих генов контрольных точек может приводить к анеуплоидии и метастазированию опухоли (38).
В то время как все больше и больше данных об этих нарушениях белков, приводящих к неправильной функции контрольных точек веретена, все еще остается открытым, является ли анеуплоидия причиной или следствием развития рака и опухолей. Аргумент в пользу анеуплоидии как следствия, а не причины рака заключается в знании того, что часто встречающиеся мутации в регуляторных путях, таких как пути p53 и pRb, приводят к геномной и хромосомной нестабильности, и что частая анеуплоидия в раковых клетках является в результате этой хромосомной нестабильности (39). Мэннинг и др. показали, что истощение pRb, которое, как обсуждалось выше, может вызывать ошибки в контрольной точке рестрикции, также приводит к ошибкам в структуре центромер, приводя к более высокой частоте меротелиального прикрепления и, следовательно, к более высокой частоте неправильной хромосомной сегрегации. В конечном итоге это может привести к аномалиям, связанным с потерей/приобретением целых хромосом (40). В этих случаях анеуплоидия возникает в результате альтернативных форм нестабильности и, хотя и влияет на развитие этих клеток, является следствием других причин.
В конечном итоге это может привести к аномалиям, связанным с потерей/приобретением целых хромосом (40). В этих случаях анеуплоидия возникает в результате альтернативных форм нестабильности и, хотя и влияет на развитие этих клеток, является следствием других причин.
С другой стороны, исследования, связанные с наблюдением числа хромосом в линиях раковых клеток, таких как трисомия определенных хромосом, показывают результирующую дупликацию мутировавших аллелей, связывающую анеуплоидию с онкогенезом (41,42). Бегини и др. предположили, что трисомия хромосомы 4 приводит к удвоению мутантного аллеля рецептора тирозинкиназы (KIT), наблюдаемому при остром миелоидном лейкозе (41), а Zhuang et al. наблюдали дупликацию другого мутантного аллеля рецептора тирозинкиназы (МЕТ), возникающего в результате трисомии хромосомы 7 при папиллярной карциноме почки (42).
В целом, особая роль, которую анеуплоидия играет в развитии рака, до сих пор неясна, а также то, способствует ли она причинам рака или является результатом других причин. Будущие исследования этих явлений должны улучшить наше понимание регуляции клеточного цикла в раковых клетках и могут указать на новые подходы к лечению рака.
Будущие исследования этих явлений должны улучшить наше понимание регуляции клеточного цикла в раковых клетках и могут указать на новые подходы к лечению рака.
Существуют различные основные механизмы, с помощью которых регулируется деление и размножение клеток. Нарушение регуляции этих механизмов может иметь пагубные и даже летальные последствия для клетки и организма. Неисправленные ошибки в клеточном цикле, включая повреждение ДНК и мутации или неправильную хромосомную сегрегацию и анеуплоидию, приводят к нестабильности генома, отличительной характеристике рака. Знание способов, с помощью которых происходит онкогенез, важно для изучения методов лечения и, в конечном итоге, излечения от рака. Обсуждение роли анеуплоидии в развитии рака указывает на область, в которой рекомендуется проводить дополнительные исследования, а также на область, которая может стать важной частью решения головоломки, объясняющей развитие рака.
Авторы заявляют об отсутствии конкурирующих финансовых интересов.
Авторы выражают благодарность Департаменту биологии Отделения естественных и социальных наук Карфагенского колледжа, Кеноша, Висконсин 53140 за поддержку в проведении исследования.
1. Моррис Дж., Хартл Д., Нолл А., Лу Р., Берри А., Бивенер А., Фаррелл Б., Холбрук Н., Пирс Н., Вил А. Биология: как устроена жизнь. Высшее образование Макмиллана, Нью-Йорк. 2013 [Google Scholar]
2. Шафер К.А. Клеточный цикл: обзор. Вет Путь. 1998;35:461–478. [PubMed] [Google Scholar]
3. Baserga R. Исследование синтеза нуклеиновых кислот в клетках асцитной опухоли методом двухэмульсионной авторадиографии. Джей Селл Биол. 1962; 12: 633–637. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Darzynkiewicz A, Gong J, Juan G, Ardelt B, Traganos F. Цитометрия циклиновых белков. Цитометрия Pt A. 1996;25(1):1–13. [Google Scholar]
5. Зеттерберг А., Ларссон О., Виман К.Г. Что является точкой ограничения. Curr Opin в Cell Biol. 1995;7(6):835–842. [PubMed] [Академия Google]
6. Сосны Дж. Циклины и циклинзависимые киназы: тема и варианты. Adv Рак Res. 1995; 66: 181–212. [PubMed] [Google Scholar]
Сосны Дж. Циклины и циклинзависимые киназы: тема и варианты. Adv Рак Res. 1995; 66: 181–212. [PubMed] [Google Scholar]
7. Малумбрес М., Барбацид М. Клеточный цикл, CDK и рак: меняющаяся парадигма. Нат Рев Рак. 2009; 9:153. [PubMed] [Google Scholar]
8. Satyanarayana A, Berthet C, Lopez-Molina J, Coppola V, Tessarollo L, Kaldis P. Генетическая замена Cdk1 на Cdk2 приводит к гибели эмбрионов и потере мейотической функции Cdk2. Разработка. 2008;135:3389–3400. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Sherr CJ, Roberts JM. Ингибиторы CDK: положительные и отрицательные регуляторы прогрессирования G1-фазы. Гены Дев. 1999; 13:1501–1512. [PubMed] [Google Scholar]
10. Кастан М.Б., Бартек Дж. Контрольные точки клеточного цикла и рак. Природа. 2004; 432:316–323. [PubMed] [Google Scholar]
11. Натарадж А.Дж., II JCT, Анантасвами Х.Н. Мутации гена p53 и фотоканцерогенез. Фотохим Фотобиол. 1995;62(2):218–230. [PubMed] [Академия Google]
12. Кларк А.Р., Гледхилл С., Хупер М.Л., Берд К.С., Уилли А.Х. Зависимость от p53 раннего апоптоза и пролиферативных ответов в эпителии кишечника мышей после гамма-облучения. Онкоген. 1994; 9 (6): 1767–1773. [PubMed] [Google Scholar]
Кларк А.Р., Гледхилл С., Хупер М.Л., Берд К.С., Уилли А.Х. Зависимость от p53 раннего апоптоза и пролиферативных ответов в эпителии кишечника мышей после гамма-облучения. Онкоген. 1994; 9 (6): 1767–1773. [PubMed] [Google Scholar]
13. Greenblatt MS, Bennett WP, Hollstein M, Harris CC. Мутации в гене супрессора опухоли p53: ключи к этиологии рака и молекулярному патогенезу. Рак Рез. 1994; 54:4855–4878. [PubMed] [Google Scholar]
14. Yao SL, Akhtar AJ, Mckenna KA, Bedi GC, Sidransky D, Mabry M, Ravi R, Collector M, Jones R, Sharkis S, Fuchs E, Bedi A. Избирательная радиосенсибилизация p53-дефицитные клетки за счет опосредованной кофеином активации киназы p34cdc2. Нат Мед. 1996;2:1140–1143. [PubMed] [Google Scholar]
15. Lavigueur AS, Maltby W, Mock D, Rossant J, Pawson T, Bernstein A. Высокая частота опухолей легких, костей и лимфоидных опухолей у трансгенных мышей со сверхэкспрессией мутантных аллелей онкогена p53. Мол Селл Биол. 1989; 9: 3982–3991. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Finlay CA. Онкоген mdm-2 может преодолевать подавление роста трансформированных клеток р53 дикого типа. Мол Селл Биол. 1993; 13: 301–306. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Finlay CA. Онкоген mdm-2 может преодолевать подавление роста трансформированных клеток р53 дикого типа. Мол Селл Биол. 1993; 13: 301–306. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Хейнс Д.С., Ландерс Дж.Е., Энгл Л.Дж., Джордж Д.Л. Физическое и функциональное взаимодействие между белками p53 и mdm2 дикого типа. Мол Селл Биол. 1994; 14:1171–1178. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Малумбрес М., Барбацид М. Циклировать или не циклировать: критическое решение при раке. Нат Рев Рак. 2001; 1: 222–231. [PubMed] [Google Scholar]
19. Arroyo M, Raychaudhuri P. Ретинобластома-репрессия E2F-зависимой транскрипции зависит от способности белка ретинобластомы взаимодействовать с E2F и отменяется онкобелком E1A аденовируса. Нуклеиновые Кислоты Res. 1992;20:5947–5954. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Zhang SY, Caamano J, Cooper F, Guo W, Klein-Szanto AJ. Иммуногистохимия циклина D1 при раке молочной железы человека. Ам Джей Клин Патол. 1994; 102: 695–698. [PubMed] [Google Scholar]
Ам Джей Клин Патол. 1994; 102: 695–698. [PubMed] [Google Scholar]
21. Джаллепалли П.В., Ленгауэр К. Хромосомная сегрегация и рак: разгадка тайны. Нат Рев Рак. 2001; 1:109. [PubMed] [Google Scholar]
22. Weaver BA, Cleveland DW. Вызывает ли анеуплоидия рак. Curr Opin Cell Biol. 2006; 18: 658–667. [PubMed] [Академия Google]
23. Paulsson K, Johansson B. Трисомия 8 как единственная хромосомная аберрация при остром миелоидном лейкозе и миелодиспластическом синдроме. Патол Биол. 2007; 55:37–48. [PubMed] [Google Scholar]
24. Thiagalingam S, Laken S, Willson JK, Markowitz SD, Kinzler KW, Vogelstein B, Lengauer C. Механизмы, лежащие в основе потери гетерозиготности при колоректальном раке человека. Proc Natl Acad Sci USA. 2001; 98: 2698–2702. [PMC free article] [PubMed] [Google Scholar]
25. Duesberg P, Rausch C, Rasnick D, Hehlmann R. Генетическая нестабильность раковых клеток пропорциональна степени их анеуплоидии. Proc Natl Acad Sci USA. 1998;95:13692–13697. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Томпсон С.Л., Комптон Д.А. Изучение связи между хромосомной нестабильностью и анеуплоидией в клетках человека. Джей Селл Биол. 2008; 180: 665–672. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Ганем Н.Дж., Годиньо С.А., Пеллман Д. Механизм, связывающий дополнительные центросомы с хромосомной нестабильностью. Природа. 2009; 460(7252):278–282. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Nasmyth K, Peters JM, Uhlmann F. Расщепление хромосомы: разрезание связей, связывающих сестринские хроматиды. Наука. 2000;288:1379–1384. [PubMed] [Google Scholar]
29. Rieder CL, Cole RW, Khodjakov A, Sluder G. Контрольная точка, задерживающая анафазу в ответ на моноориентацию хромосом, опосредована ингибирующим сигналом, производимым неприкрепленными кинетохорами. Джей Селл Биол. 1995; 130:941–948. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Shah JV, Cleveland DW. В ожидании анафазы: Mad2 и контрольная точка сборки шпинделя. Клетка. 2000;103:997–1000. [PubMed] [Google Scholar]
31. Cahill DP, Lengauer C, Yu J, Riggins GJ, Willson JK, Markowitz SD, Kinzler KW и Vogelstein B. Мутации генов митотических контрольных точек при раке человека. Природа. 1998;392:300–303. [PubMed] [Google Scholar]
32. Fang G, Yu H, Kirschner MW. Прямое связывание членов семейства белков CDC20 активирует комплекс, стимулирующий анафазу, в митозе и G1. Мол Ячейка. 1998; 2: 163–171. [PubMed] [Google Scholar]
33. Бхарадвадж Р., Ю. Х. Контрольная точка веретена, анеуплоидия и рак. Онкоген. 2004;23:2016–2027. [PubMed] [Google Scholar]
34. Lengauer C, Kinzler KW, Vogelstein B. Генетическая нестабильность при колоректальном раке. Природа. 1997; 386: 623–627. [PubMed] [Академия Google]
35. Li Y, Benezra R. Идентификация гена митотической контрольной точки человека: hsMAD2. Наука. 1996; 247: 246–248. [PubMed] [Google Scholar]
36. Gemma A, Seike Y, Uematsu K, Hibino S, Kurimoto F, Yoshimura A, Shibuya M, Harris C, Kudoh S. Соматическая мутация hBUB1 гена митотической контрольной точки в первичной рак легких. Гены Хромосомы Рак. 2000; 29: 213–218. [PubMed] [Google Scholar]
37. Ohshima K, Haraoka S, Yoshioka S, Hamasaki M, Fujiki T, Suzumiya J, Kawasaki C, Kanda M, Kikuchi M. Мутационный анализ генов митотических контрольных точек ( hBUB1 и hBUBR1 ) и микросателлитной нестабильности при Т-клеточном лейкозе/лимфоме взрослых. Рак Летт. 2000; 158:141–150. [PubMed] [Google Scholar]
38. Shichiri M, Yoshinago K, Hisatomi H, Sugihara K, Hirata Y. Генетическая и эпигенетическая инактивация генов митотических контрольных точек hBUB1 и hBUBR1 и их связь с выживанием. Рак Рез. 2002; 62:13–17. [PubMed] [Google Scholar]
39. Gordon DJ, Resio B, Pellman D. Причины и последствия анеуплоидии при раке. Нат Рев Жене. 2012;13:189. [PubMed] [Google Scholar]
40. Manning AL, Longworth MS, Dyson NJ. Потеря pRB вызывает дисфункцию центромер и хромосомную нестабильность. Гены Дев. 2010; 24:1364–1376. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Beghini A, Ripamonti CB, Castorina P, Pezzetti L, Doneda L, Cairoli R, Morra E, Larizza L. Трисомия 4, приводящая к дублированию мутировавшего аллеля KIT при остром миелоидном лейкозе с поражением тучных клеток. Рак Генет Цитогенет. 2000; 119: 26–31. [PubMed] [Академия Google]
42. Zhuang Z, Park WS, Pack S, Schmidt L, Vortmeyer AO, Pak E, Pham T, Weil R, Sonja C, Lubensky I, Linehan W, Zbar B, Weirich G. Трисомия 7-укрывающая неслучайная удвоение мутантного аллеля MET при наследственном папиллярном раке почки. Нат Жене. 1998; 20:66–69. [PubMed] [Google Scholar]
контрольных точек клеточного цикла | Биология для специальностей I
Определите и объясните важные контрольные точки, через которые клетка проходит в течение клеточного цикла
Как мы только что узнали, клеточный цикл — довольно сложный процесс. Чтобы убедиться, что все идет правильно, в цикле есть контрольные точки. Давайте узнаем об этом и о том, как они помогают контролировать клеточный цикл.
Цели обучения
- Определение важных контрольных точек в делении клеток
- Объясните, как ошибки в клеточном делении связаны с раком
Продолжительность клеточного цикла сильно варьирует даже в клетках одного организма. У людей частота обновления клеток колеблется от нескольких часов в раннем эмбриональном развитии до в среднем от двух до пяти дней для эпителиальных клеток и до всей жизни человека, проводимой в G 0 специализированными клетками, такими как нейроны коры. или клеток сердечной мышцы. Также различается время, которое клетка проводит в каждой фазе клеточного цикла. При выращивании быстроделящихся клеток млекопитающих в культуре (вне организма при оптимальных условиях выращивания) продолжительность цикла составляет около 24 часов. В быстро делящихся клетках человека с 24-часовым клеточным циклом G 9Фаза 0017 1 длится примерно девять часов, фаза S длится 10 часов, фаза G 2 длится около четырех с половиной часов, а фаза М длится примерно полчаса. У ранних эмбрионов плодовых мушек клеточный цикл завершается примерно за восемь минут. Время событий в клеточном цикле контролируется механизмами, которые являются как внутренними, так и внешними по отношению к клетке.
Регулирование клеточного цикла внешними событиями
Как инициация, так и ингибирование клеточного деления запускаются внешними по отношению к клетке событиями, когда она собирается начать процесс репликации. Событие может быть таким простым, как гибель соседней клетки, или столь масштабным, как высвобождение гормонов, стимулирующих рост, таких как гормон роста человека (HGH). Недостаток гормона роста может подавлять деление клеток, что приводит к карликовости, тогда как избыток гормона роста может привести к гигантизму. Скученность клеток также может препятствовать делению клеток. Другим фактором, который может инициировать клеточное деление, является размер клетки; по мере роста клетки она становится неэффективной из-за уменьшающегося отношения поверхности к объему. Решение этой проблемы — разделить.
Каким бы ни был источник сообщения, ячейка получает сигнал, и ряд событий внутри ячейки позволяет ему перейти в интерфазу. Двигаясь вперед от этой точки инициации, каждый параметр, требуемый на каждой фазе клеточного цикла, должен быть соблюден, иначе цикл не может развиваться.
Регулирование на внутренних контрольно-пропускных пунктах
Очень важно, чтобы полученные дочерние клетки были точными копиями родительской клетки. Ошибки в удвоении или распределении хромосом приводят к мутациям, которые могут передаваться каждой новой клетке, полученной из аномальной клетки. Чтобы предотвратить дальнейшее деление скомпрометированной клетки, существуют механизмы внутреннего контроля, которые работают на трех основных контрольных точках клеточного цикла. Контрольная точка — это одна из нескольких точек в эукариотическом клеточном цикле, в которой продвижение клетки к следующей стадии цикла может быть остановлено до тех пор, пока условия не станут благоприятными. Эти контрольные точки происходят ближе к концу G 1 , при переходе G 2 /M и во время метафазы (рис. 1).
Рисунок 1. Клеточный цикл контролируется в трех контрольных точках. Целостность ДНК оценивается на контрольно-пропускном пункте G 1 . Правильная дупликация хромосом оценивается в контрольной точке G 2 . Прикрепление каждой кинетохоры к волокну веретена оценивают в контрольной точке М.
Контрольная точка G
1 Контрольная точка G 1 определяет, все ли условия благоприятны для продолжения деления клеток. G 1 Контрольная точка, также называемая точкой рестрикции (у дрожжей), представляет собой точку, в которой клетка необратимо вступает в процесс клеточного деления. Внешние воздействия, такие как факторы роста, играют большую роль в переносе клетки через контрольную точку G 1 . В дополнение к адекватным запасам и размеру клеток, существует проверка на наличие повреждений геномной ДНК на контрольно-пропускном пункте G 1 . Ячейка, которая не соответствует всем требованиям, не будет допущена к этапу S. Клетка может остановить цикл и попытаться исправить проблемное состояние или перейти в G9.0017 0 и ждите дальнейших сигналов, когда условия улучшатся.
Контрольная точка G
2Контрольная точка G 2 блокирует вступление в митотическую фазу, если не выполняются определенные условия. Как и в контрольной точке G 1 , оценивают размер клеток и запасы белка. Однако наиболее важная роль контрольной точки G 2 заключается в том, чтобы гарантировать, что все хромосомы были реплицированы и что реплицированная ДНК не повреждена. Если механизмы контрольных точек обнаруживают проблемы с ДНК, клеточный цикл останавливается, и клетка пытается либо завершить репликацию ДНК, либо восстановить поврежденную ДНК.
Контрольная точка M
Контрольная точка M возникает в конце метафазной стадии кариокинеза. Контрольная точка М также известна как контрольная точка веретена, потому что она определяет, правильно ли все сестринские хроматиды прикреплены к микротрубочкам веретена. Поскольку разделение сестринских хроматид во время анафазы является необратимым этапом, цикл не будет продолжаться до тех пор, пока кинетохоры каждой пары сестринских хроматид не будут прочно прикреплены по крайней мере к двум волокнам веретена, отходящим от противоположных полюсов клетки.
Посмотрите, что происходит на контрольных точках G 1 , G 2 и M, загрузив эту анимацию клеточного цикла.
Молекулы-регуляторы клеточного цикла
Помимо внутренне контролируемых контрольных точек, существуют две группы внутриклеточных молекул, регулирующих клеточный цикл. Эти регуляторные молекулы либо способствуют переходу клетки к следующей фазе (позитивная регуляция), либо останавливают цикл (негативная регуляция). Молекулы-регуляторы могут действовать индивидуально или влиять на активность или продукцию других регуляторных белков. Следовательно, выход из строя одного регулятора может почти не влиять на клеточный цикл, особенно если одно и то же событие контролируется более чем одним механизмом. И наоборот, эффект недостаточного или нефункционирующего регулятора может быть широким и, возможно, фатальным для клетки, если затрагиваются несколько процессов.
Положительная регуляция клеточного цикла
Две группы белков, называемых циклинами и циклин-зависимыми киназами (Cdks), отвечают за продвижение клетки через различные контрольные точки. Уровни четырех циклиновых белков колеблются на протяжении клеточного цикла по предсказуемой схеме (рис. 2). Повышение концентрации белков циклинов вызывается как внешними, так и внутренними сигналами. После перехода клетки на следующую стадию клеточного цикла происходит деградация циклинов, которые были активны на предыдущей стадии.
Рисунок 2. Концентрации белков циклинов изменяются на протяжении клеточного цикла. Существует прямая корреляция между накоплением циклина и тремя основными контрольными точками клеточного цикла. Также обратите внимание на резкое снижение уровня циклина после каждой контрольной точки (переход между фазами клеточного цикла), поскольку циклин расщепляется цитоплазматическими ферментами. (кредит: модификация работы «WikiMiMa»/Wikimedia Commons)
Циклины регулируют клеточный цикл только тогда, когда они тесно связаны с Cdks. Чтобы быть полностью активным, комплекс Cdk/циклин также должен быть фосфорилирован в определенных местах. Как и все киназы, Cdks представляют собой ферменты (киназы), которые фосфорилируют другие белки. Фосфорилирование активирует белок, изменяя его форму. Белки, фосфорилированные Cdks, участвуют в переходе клетки к следующей фазе (рис. 3). Уровни белков Cdk относительно стабильны на протяжении клеточного цикла; однако концентрации циклина колеблются и определяют, когда образуются комплексы Cdk/циклин. Различные циклины и Cdks связываются в определенных точках клеточного цикла и, таким образом, регулируют различные контрольные точки.
Рисунок 3. Циклинзависимые киназы (Cdks) — это протеинкиназы, которые при полной активации могут фосфорилировать и, таким образом, активировать другие белки, которые продвигают клеточный цикл после контрольной точки. Чтобы полностью активироваться, Cdk должен связываться с белком циклином, а затем фосфорилироваться другой киназой.
Поскольку циклические колебания уровней циклина основаны на времени клеточного цикла, а не на конкретных событиях, регуляция клеточного цикла обычно происходит либо только молекулами Cdk, либо комплексами Cdk/циклин. Без определенной концентрации полностью активированных комплексов циклин/Cdk клеточный цикл не может проходить через контрольные точки.
Хотя циклины являются основными регуляторными молекулами, определяющими поступательный импульс клеточного цикла, существует несколько других механизмов, которые точно настраивают ход цикла с негативными, а не позитивными эффектами. Эти механизмы по существу блокируют развитие клеточного цикла до тех пор, пока проблемные состояния не будут устранены. Молекулы, препятствующие полной активации Cdk, называются ингибиторами Cdk. Многие из этих молекул-ингибиторов прямо или косвенно контролируют конкретное событие клеточного цикла. Блокировка, помещенная на Cdks молекулами ингибитора, не будет удалена до тех пор, пока не завершится конкретное событие, которое отслеживает ингибитор.
Негативная регуляция клеточного цикла
Вторая группа молекул, регулирующих клеточный цикл, представляет собой негативные регуляторы. Отрицательные регуляторы останавливают клеточный цикл. Помните, что при позитивной регуляции активные молекулы вызывают прогресс цикла.
Наиболее изученными негативными регуляторными молекулами являются белок ретинобластомы (Rb), p53 и p21. Белки ретинобластомы представляют собой группу белков-супрессоров опухолей, распространенных во многих клетках. Обозначения 53 и 21 относятся к функциональной молекулярной массе белков (p) в килодальтонах. Многое из того, что известно о регуляции клеточного цикла, получено из исследований, проведенных с клетками, утратившими регулирующий контроль. Было обнаружено, что все три из этих регуляторных белков повреждены или нефункциональны в клетках, которые начали бесконтрольно реплицироваться (стали раковыми). В каждом случае основной причиной неконтролируемого прохождения клеточного цикла была дефектная копия регуляторного белка.
Rb, p53 и p21 действуют преимущественно на КПП G 1 . р53 является многофункциональным белком, который оказывает большое влияние на готовность клетки к делению, поскольку он действует, когда в клетках, находящихся в процессе подготовки к G 1 , имеется поврежденная ДНК. Если обнаруживается поврежденная ДНК, p53 останавливает клеточный цикл и привлекает ферменты для восстановления ДНК. Если ДНК не может быть восстановлена, p53 может вызвать апоптоз или самоубийство клеток, чтобы предотвратить дублирование поврежденных хромосом. По мере повышения уровня p53 запускается производство p21. p21 обеспечивает остановку цикла, продиктованного p53, путем связывания и ингибирования активности комплексов Cdk/циклин. По мере того, как клетка подвергается большему стрессу, накапливаются более высокие уровни p53 и p21, что снижает вероятность того, что клетка перейдет в S-фазу.
Rb оказывает регулирующее влияние на другие белки-положительные регуляторы. Главным образом, Rb контролирует размер клеток. В активном дефосфорилированном состоянии Rb связывается с белками, называемыми факторами транскрипции, чаще всего с E2F (рис. 4). Факторы транскрипции «включают» определенные гены, позволяя производить белки, кодируемые этим геном. Когда Rb связан с E2F, блокируется продукция белков, необходимых для перехода G 1 /S. По мере увеличения размера клетки Rb медленно фосфорилируется, пока не станет инактивированным. Rb высвобождает E2F, который теперь может включать ген, производящий переходный белок, и этот конкретный блок снимается. Чтобы ячейка прошла каждую из контрольных точек, все положительные регуляторы должны быть «включены», а все отрицательные регуляторы должны быть «выключены».
Практический вопрос
Рисунок 4. Rb останавливает клеточный цикл и освобождает его от удержания в ответ на рост клеток.
Rb и другие белки, негативно регулирующие клеточный цикл, иногда называют супрессорами опухолей. Как вы думаете, почему для этих белков подходит название «опухолесупрессор»?
Показать ответ
Рак и клеточный цикл
Рак включает множество различных заболеваний, вызываемых общим механизмом: неконтролируемым ростом клеток. Несмотря на избыточность и перекрывающиеся уровни контроля клеточного цикла, ошибки случаются. Одним из критических процессов, отслеживаемых механизмом наблюдения за контрольными точками клеточного цикла, является правильная репликация ДНК во время S-фазы. Даже когда все элементы управления клеточным циклом полностью функциональны, небольшой процент ошибок репликации (мутаций) будет передан дочерним клеткам. Если изменения в последовательности нуклеотидов ДНК происходят в кодирующей части гена и не исправляются, возникает мутация гена. Все виды рака начинаются, когда генная мутация приводит к дефектному белку, который играет ключевую роль в репродукции клеток. Изменения в клетке, возникающие в результате деформированного белка, могут быть незначительными: возможно, небольшая задержка в связывании Cdk с циклином или с белком Rb, который отсоединяется от своей ДНК-мишени, оставаясь при этом фосфорилированным. Однако даже незначительные ошибки могут способствовать более легкому совершению последующих ошибок. Снова и снова небольшие неисправленные ошибки передаются от родительской клетки к дочерним клеткам и усиливаются по мере того, как каждое поколение производит больше нефункциональных белков из неисправленных повреждений ДНК. В конце концов, скорость клеточного цикла увеличивается по мере снижения эффективности механизмов контроля и восстановления. Неконтролируемый рост мутировавших клеток опережает рост нормальных клеток в этой области, и опухоль (~ oma ).
Протоонкогены
Гены, кодирующие регуляторы положительного клеточного цикла, называются протоонкогенами . Протоонкогены — это нормальные гены, которые при определенных мутациях становятся онкогенами , генами, вызывающими превращение клетки в раковую. Рассмотрим, что может произойти с клеточным циклом в клетке с недавно приобретенным онкогеном. В большинстве случаев изменение последовательности ДНК приводит к менее функциональному (или нефункциональному) белку. Результат вреден для клетки и, вероятно, помешает клетке завершить клеточный цикл; однако организму не наносится вред, потому что мутация не будет перенесена. Если клетка не может воспроизводиться, мутация не распространяется и ущерб минимален. Однако иногда мутация гена вызывает изменение, которое увеличивает активность положительного регулятора. Например, мутация, которая позволяет активировать Cdk без взаимодействия с циклином, может подтолкнуть клеточный цикл к контрольной точке до того, как будут выполнены все необходимые условия. Если полученные дочерние клетки будут слишком повреждены, чтобы подвергаться дальнейшим клеточным делениям, мутация не будет распространяться, и организму не будет нанесено никакого вреда. Однако, если атипичные дочерние клетки способны подвергаться дальнейшим клеточным делениям, последующие поколения клеток, вероятно, будут накапливать еще больше мутаций, некоторые, возможно, в дополнительных генах, регулирующих клеточный цикл.
Ген Cdk в приведенном выше примере является лишь одним из многих генов, которые считаются протоонкогенами. В дополнение к белкам, регулирующим клеточный цикл, любой белок, влияющий на цикл, может быть изменен таким образом, чтобы перекрыть контрольные точки клеточного цикла. Онкоген — это любой ген, изменение которого приводит к увеличению скорости прогрессирования клеточного цикла.
Гены-супрессоры опухолей
Подобно протоонкогенам, многие белки, регулирующие негативный клеточный цикл, были обнаружены в клетках, ставших раковыми. Гены-супрессоры опухолей представляют собой сегменты ДНК, которые кодируют негативные регуляторные белки, тип регуляторов, которые при активации могут предотвратить неконтролируемое деление клетки. Коллективная функция наиболее изученных белков генов-супрессоров опухолей, Rb, p53 и p21, состоит в том, чтобы блокировать развитие клеточного цикла до завершения определенных событий. Клетка, несущая мутированную форму негативного регулятора, может быть не в состоянии остановить клеточный цикл, если возникнет проблема. Подавители опухолей подобны тормозам в автомобиле: неисправность тормозов может привести к автокатастрофе.
Мутированные гены p53 были идентифицированы более чем в половине всех опухолевых клеток человека. Это открытие неудивительно в свете многочисленных ролей, которые белок р53 играет в контрольной точке G 1 . Клетка с неисправным р53 может не обнаружить ошибки, присутствующие в геномной ДНК (рис. 5). Даже если частично функциональный p53 действительно идентифицирует мутации, он больше не может сигнализировать о необходимых ферментах репарации ДНК. В любом случае поврежденная ДНК останется неисправленной. В этот момент функциональный р53 сочтет клетку нежизнеспособной и вызовет запрограммированную гибель клетки (апоптоз). Однако поврежденная версия p53, обнаруженная в раковых клетках, не может запускать апоптоз.
Рисунок 5. Роль нормального р53 заключается в мониторинге ДНК и снабжении кислородом (гипоксия — это состояние пониженного снабжения кислородом). При обнаружении повреждения p53 запускает механизмы восстановления. Если восстановление не удается, р53 сигнализирует об апоптозе. Клетка с аномальным белком p53 не может восстанавливать поврежденную ДНК и, следовательно, не может сигнализировать об апоптозе. Клетки с аномальным p53 могут стать раковыми. (кредит: модификация работы Thierry Soussi)
Потеря функции p53 имеет и другие последствия для клеточного цикла. Мутированный p53 может потерять способность запускать продукцию p21. Без адекватных уровней p21 не существует эффективного блока активации Cdk. По сути, без полностью функционального p53 G 9Контрольная точка 0017 1 серьезно нарушена, и ячейка переходит непосредственно из G 1 в S независимо от внутренних и внешних условий. По завершении этого укороченного клеточного цикла образуются две дочерние клетки, унаследовавшие мутировавший ген p53. Учитывая неоптимальные условия, в которых воспроизводилась родительская клетка, вполне вероятно, что дочерние клетки приобретут другие мутации в дополнение к неисправному гену-супрессору опухоли. Такие клетки, как эти дочерние клетки, быстро накапливают как онкогены, так и нефункциональные гены-супрессоры опухолей. Опять же, результатом является рост опухоли.
В этом видео рассказывается о том, что рак является побочным продуктом нарушенной репликации ДНК:
Вкратце: Контрольные точки клеточного цикла
Каждый этап клеточного цикла контролируется внутренним контролем, называемым контрольными точками. В клеточном цикле есть три основных контрольных точки: одна в конце G 1 , вторая в переходе G 2 /M и третья во время метафазы. Положительные регуляторные молекулы позволяют клеточному циклу перейти на следующую стадию. Молекулы отрицательного регулятора контролируют клеточное состояние и могут останавливать цикл до тех пор, пока не будут выполнены определенные требования.
Рак является результатом бесконтрольного деления клеток, вызванного нарушением механизмов, регулирующих клеточный цикл. Потеря контроля начинается с изменения последовательности ДНК гена, кодирующего одну из регуляторных молекул.