Характеристика кия рио: Технические характеристики Kia Rio 2021
KIA Rio 1.4 MT: цена, технические характеристики КИА Рио 1.4 MT
KIA Rio 1.4 MT: цена, технические характеристики КИА Рио 1.4 MT — Avto-Russia.ru- Главная
- Каталог авто
- KIA
- KIA Rio
- KIA Rio 1.4 MT
Поиск по каталогу
Тип кузова: Любой Седан Хэтчбек Универсал Кроссовер Внедорожник Компактвэн Минивэн Купе Кабриолет Родстер Пикап Фургон Автобус Микроавтобус Грузовик Самосвал Шасси ТягачДиапазон цен: Любой до 500 000 руб от 500 000 до 600 000 руб от 500 000 до 600 000 руб от 600 000 до 700 000 руб от 700 000 до 800 000 руб от 800 000 до 900 000 руб от 900 000 до 1 000 000 руб до 1 000 000 руб от 1 250 000 до 1 500 000 руб от 1 250 000 до 1 500 000 руб от 1 500 000 до 1 750 000 руб от 1 750 000 до 2 000 000 руб до 2 000 000 руб от 2 000 000 до 2 500 000 руб от 2 500 000 до 3 000 000 руб от 3 000 000 до 3 500 000 руб от 3 500 000 до 4 000 000 руб от 4 000 000 до 4 500 000 руб от 4 500 000 до 5 000 000 руб свыше 5 000 000 рубДлина: Любая До 3 метров 3 — 3,5 метра 3,5 — 4 метра 4 — 4,5 метра 4,5 — 5 метров 5 — 5,5 метра 5,5 — 6 метров Свыше 6 метровШирина
От официальных дилеров
Модели 2022 года
Исключить китайские авто
Поиск Все марки
- Фото
- Модификации
- Одноклассники
- Отзывы
- Обои
Основные характеристики
| Марка | KIA |
| Модель | KIA Rio |
| Модификация | KIA Rio 1. 4 MT 4 MT |
| Модельный год | 2017 |
| Тип кузова | Седан |
| Количество дверей | 4 |
| Количество мест | 5 |
| Страна сборки | Россия |
| Гарантия | 5 лет или 150 000 км |
| Цена (январь 2022 г) | 805 000 ₽ |
Эксплуатационные характеристики
| Вид топлива | АИ-92 |
| Время разгона до 100 км/ч | 12.2 сек |
| Максимальная скорость | |
| Расход топлива в городском цикле | 7.2 л на 100 км |
| Расход топлива на трассе | 4.8 л на 100 км |
| Расход топлива в смешанном цикле | 5.7 л на 100 км |
| Запас хода | 694 — 1 042 км |
| Расходы на топливо в год (при пробеге 100 км в день) | 98 824 ₽ |
| Транспортный налог * (Москва) | 1 200 ₽ |
| ОСАГО * (Москва, возраст свыше 22 лет, стаж более 3 лет) | 7 570 ₽ |
| КАСКО (Москва, возраст свыше 22 лет, стаж более 3 лет) | 28 175 — 68 425 ₽ |
* Воспользуйтесь калькуляторами Налога и ОСАГО для более детального расчета.
Габариты и размеры
| Длина | 4400 мм |
| Ширина | 1740 мм |
| Высота | 1470 мм |
| Дорожный просвет | 160 мм |
| Колея передняя | 1516 мм |
| Колея задняя | 1524 мм |
| Колесная база | 2600 мм |
Масса
| Снаряженная масса | 1150 кг |
| Полная масса | 1560 кг |
| Грузоподъемность | 410 кг |
Объемы
| Объем багажника | 480 л |
| Объем топливного бака | 50 л |
Двигатель
| Тип двигателя | Бензиновый |
| Число цилиндров / расположение | 4/Рядный |
Мощность двигателя, л. с / оборотах с / оборотах | 100/6000 |
| Рабочий объем двигателя | 1368 см³ |
| Крутящий момент, Н·м / оборотах | 132/4000 |
Трансмиссия
| Привод | Передний |
| Тип коробки передач | Механическая, 6 передач |
Руль
| Усилитель руля | Электроусилитель |
Электронные системы
| Электронные системы управления | ABS,EBA,EBD,ESP,HHC |
Климат
| Управление климатом | Кондиционер |
Подвеска
| Передняя подвеска | Независимая, McPherson |
| Задняя подвеска | Полузависимая, торсионная балка |
Тормоза
| Передние тормоза | Дисковые, вентилируемые |
| Задние тормоза | Барабанные |
Шины и диски
| Размер шин | 185/65 R15 |
| Размер дисков | 15x6J |
- Фото
- Модификации
- Одноклассники
- Отзывы
- Обои
Седан KIA Rio
Сообщить об ошибке
Технические характеристики КИА Рио Х Лайн
Технические характеристики КИА Рио Х Лайн показаны в таблицах – габаритные размеры, подвеска, двигатели, коробки передачи и т. д.
д.
Содержание
Габаритные размеры (мм)
| Длина | 4 240 |
| Ширина | 1 750 |
| Высота | 1 535 |
| Колесная база | 2 600 |
| Колея передних колес | 1 507 — 1 513 |
| Колея задних колес | 1 524 — 1 530 |
| Передний свес | 845 |
| Задний свес | 795 |
| Дорожный просвет | 195 мм с колесами R16, 190 мм с колесами R15 |
Внутренние размеры (мм)
| Пространство для ног (1-й/2-й/3-й ряд) | 1 070/870 |
| Расстояние от подушки сидения до потолка (1-й/2-й/3-й ряд) | 991/966 |
| Ширина салона на уровне плеч (1-й/2-й/3-й ряд) | 1 375/1 365 |
| Объем бачка омывающей жидкости (л) | 4.6 |
Вместимость
| Объем багажника (л) (VDA) | 390 |
| Объем багажника при сложенных задних сиденьях (л) | 1 075 |
Двигатели
| Тип двигателя | Бензиновый | |
| Технология двигателя | MPI | |
| Объем двигателя (л) | 1. 4 4 | 1.6 |
| Максимальная мощность (л. с.) | 99.6 | 123.0 |
| Максимальная мощность (об./мин) | 6 000 | 6 300 |
| Максимальный крутящий момент (Нм) | 132.0 | 151.0 |
| Максимальный крутящий момент (об./мин) | 4 000 | 4 850 |
| Рабочий объем (см³) | 1 368 | 1 591 |
| Диаметр цилиндра х Ход поршня (мм) | 72.0 х 84.0 | 77.0 х 85.4 |
| Степень сжатия | 10.5 | |
| Максимальная мощность (КВт (об/мин) | 73.3/6 000 | 90.2/6 300 |
| Газораспределительный механизм | 16 клапанный, с системой регулировки фаз газораспределения впускных и выпускных клапанов (D-CVVT) | |
| Топливная система | Распределенный впрыск топлива с электронным управлением | |
| Требования к топливу | Неэтилированный бензин с октановым числом не менее 92 | |
| Объем масла в двигателе (л) | 3. 3 3 | |
| Количество цилиндров и расположение | 4, Рядное | |
| Экологический класс | Евро-5 | |
Трансмиссия
| Двигатель | 1.4 (100 л. с.) | 1.4 (100 л. с.) | 1.6 (123 л. с.) | 1.6 (123 л. с.) |
| Тип привода | Передний | |||
| Тип трансмиссии | МТ | АТ | МТ | АТ |
| Количество передач | 6 | |||
| Тип сцепления | Сухое, однодисковое | Гидравлическая муфта | Сухое, однодисковое | Гидравлическая муфта |
| Объем масла в трансмиссии (л) | 1.6-1.7 | 6.7 | 1.6-1.7 | 6.7 |
Динамика
| Двигатель | 1. 4 (100 л. с.) 4 (100 л. с.) | 1.4 (100 л. с.) | 1.6 (123 л. с.) | 1.6 (123 л. с.) |
| Разгон 0 — 100 км/ч | 12.6 | 13.4 | 10.7 | 11.6 |
| Разгон 80 — 120 км/ч | 16.9 | 10.1 | 15.0 | 8.7 |
| Максимальная скорость км/ч | 176 | 174 | 184 | 183 |
Топливная экономичность
| Двигатель | 1.4 (100 л. с.) | 1.4 (100 л. с.) | 1.6 (123 л. с.) | 1.6 (123 л. с.) |
| Расход топлива (л/100 км) Комбинированный | 5.9 | 6.6 | 6.8 | |
| Город (л/100 км) | 7.4 | 8.6 | 8.7 | 8.9 |
| Трасса (л/100 км) | 5.0 | 5.4 | 5.6 | |
| Объем топливного бака (л) | 50 | |||
Рулевое управление
| Тип рулевого управления | Рулевой привод с электроусилителем, тип: шестерня-рейка |
| Передаточное число рулевого управления | 14. 6:1 6:1 |
| Число оборотов руля между крайними положениями | 2.70 |
| Минимальный радиус поворота (м_ | 5.2 |
Подвеска
| Передняя | Независимая, пружинная, типа Макферсон, с гидравлическими телескопическими амортизаторами, со стабилизатором поперечной устойчивости |
| Задняя | Полузависимая, пружинная, с гидравлическими телескопическими амортизаторами |
Масса (кг)
| Двигатель | 1.4 (100 л. с.) | 1.4 (100 л. с.) | 1.6 (123 л. с.) | 1.6 (123 л. с.) |
| Снаряженная минимальная | 1 155 | 1 187 | 1 175 | 1 203 |
| Снаряженная максимальная | 1 221 | 1 253 | 1 241 | 1 269 |
| Полная масса | 1 570 | 1 610 | 1 590 | 1 620 |
Тормозная система
| Передние тормоза | Дисковые вентилируемые |
| Задние тормоза | Дисковые (барабанные) |
Вакуумный усилитель тормозов. Передаточное число напорного усилителя Передаточное число напорного усилителя | 8:1 |
| Главный тормозной цилиндр, Тип | Фиксированный |
Электрооборудование
| Двигатель | 1.4 (100 л. с.) | 1.4 (100 л. с.) | 1.6 (123 л. с.) | 1.6 (123 л. с.) |
| Емкость аккумулятора (А/ч) | 60 | |||
| Генератор (В, А) | 13.5, 90 | 13.5, 120 | ||
| Стартер (В, КВт) | 12, 0.9 | |||
Из таблиц видно, что технические характеристики КИА Рио Х Лайн вполне оправдывают цену автомобиля.
FacebookTwitterRedditPinterestGoogle+LinkedInStumbleUponvKontakte
Kia Rio X – технические характеристики Киа Rio X 2021/2022 по цене от 1574900 руб
Двигатель и трансмиссия
1. 4 MPI
4 MPI
1.6 MPI
1.4 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
100
123
100
123
123
123
123
123
123
72.0 х 84.0
77.0 х 85.4
72.0 х 84.0
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
передний
передний
передний
передний
передний
передний
передний
передний
передний
10.5
10.5
10.5
10.5
10.5
10.5
10.5
10.5
10.5
132
151
132
151
151
151
151
151
151
100 @ 6000
123 @ 6300
100 @ 6000
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
132 @ 4000
151 @ 4850
132 @ 4000
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
1. 4
4
1.6
1.4
1.6
1.6
1.6
1.6
1.6
1.6
1368
1591
1368
1591
1591
1591
1591
1591
1591
4
4
4
4
4
4
4
4
4
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Механика (6MT)
Механика (6MT)
Автомат (6AT)
Автомат (6AT)
Механика (6MT)
Автомат (6AT)
Автомат (6AT)
Автомат (6AT)
Автомат (6AT)
передний
передний
передний
передний
передний
передний
передний
передний
передний
12,6
10,7
13,4
11,6
10,7
11,6
10,7
11,6
11,6
5,9
6,6
6,6
6,8
6,6
6,8
6,6
6,8
6,8
Сухое однодисковое
Сухое однодисковое
Гидравлическая муфта
Гидравлическая муфта
Сухое однодисковое
Гидравлическая муфта
Сухое однодисковое
Гидравлическая муфта
Гидравлическая муфта
1. 6~1.7
6~1.7
1.6~1.7
6.7
6.7
1.6~1.7
6.7
1.6~1.7
6.7
6.7
Выбросы CO2
175
202
200
207
202
207
202
207
207
117
126
126
130
126
130
126
130
130
140
154
154
158
154
158
154
158
158
Размеры
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
2600
2600
2600
2600
2600
2600
2600
2600
2600
190
190
190
190
190
190
190
190
195
390
390
390
390
390
390
390
390
390
Электрооборудование
60
60
60
60
60
60
60
60
60
13. 5 В 90 А
5 В 90 А
13.5 В 120 А
13.5 В 90 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
3.5
3.6
3.5
3.6
3.6
3.6
3.6
3.6
3.6
Рулевое управление
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
2.7
2.7
2.7
2.7
2. 7
7
2.7
2.7
2.7
2.7
5.2
5.2
5.2
5.2
5.2
5.2
5.2
5.2
5.2
Тормоза
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
Внутренние размеры
390
390
390
390
390
390
390
390
390
1075
1075
1075
1075
1075
1075
1075
1075
1075
Динамические характеристики
175
183
173
182
183
182
183
182
182
12.6
10.7
13.4
11.6
10.7
11.6
10.7
11.6
11.6
17.0
15.2
10.2
8.8
15.2
8.8
15.2
8.8
8.8
Масса (5 мест)
1570
1590
1610
1620
1590
1620
1590
1620
1620
Расход топлива
50
50
50
50
50
50
50
50
50
Спецификация
H0S6K4617
H0S6D2617
H0S6K461F
H0S6D261F
H0S6D2617
H0S6D261F
H0S6D261F
H0S6D261F
H0S6D261F
Kia Rio X – технические характеристики Киа Rio X 2021/2022 по цене от 1574900 руб
Двигатель и трансмиссия
1. 4 MPI
4 MPI
1.6 MPI
1.4 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
100
123
100
123
123
123
123
123
123
72.0 х 84.0
77.0 х 85.4
72.0 х 84.0
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
передний
передний
передний
передний
передний
передний
передний
передний
передний
10.5
10.5
10.5
10.5
10.5
10.5
10.5
10.5
10.5
132
151
132
151
151
151
151
151
151
100 @ 6000
123 @ 6300
100 @ 6000
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
132 @ 4000
151 @ 4850
132 @ 4000
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
1. 4
4
1.6
1.4
1.6
1.6
1.6
1.6
1.6
1.6
1368
1591
1368
1591
1591
1591
1591
1591
1591
4
4
4
4
4
4
4
4
4
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Механика (6MT)
Механика (6MT)
Автомат (6AT)
Автомат (6AT)
Механика (6MT)
Автомат (6AT)
Автомат (6AT)
Автомат (6AT)
Автомат (6AT)
передний
передний
передний
передний
передний
передний
передний
передний
передний
12,6
10,7
13,4
11,6
10,7
11,6
10,7
11,6
11,6
5,9
6,6
6,6
6,8
6,6
6,8
6,6
6,8
6,8
Сухое однодисковое
Сухое однодисковое
Гидравлическая муфта
Гидравлическая муфта
Сухое однодисковое
Гидравлическая муфта
Сухое однодисковое
Гидравлическая муфта
Гидравлическая муфта
1. 6~1.7
6~1.7
1.6~1.7
6.7
6.7
1.6~1.7
6.7
1.6~1.7
6.7
6.7
Выбросы CO2
175
202
200
207
202
207
202
207
207
117
126
126
130
126
130
126
130
130
140
154
154
158
154
158
154
158
158
Размеры
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
2600
2600
2600
2600
2600
2600
2600
2600
2600
190
190
190
190
190
190
190
190
195
390
390
390
390
390
390
390
390
390
Электрооборудование
60
60
60
60
60
60
60
60
60
13. 5 В 90 А
13.5 В 120 А
13.5 В 90 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
3.5
3.6
3.5
3.6
3.6
3.6
3.6
3.6
3.6
Рулевое управление
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
2.7
2.7
2.7
2.7
2. 7
7
2.7
2.7
2.7
2.7
5.2
5.2
5.2
5.2
5.2
5.2
5.2
5.2
5.2
Тормоза
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
Внутренние размеры
390
390
390
390
390
390
390
390
390
1075
1075
1075
1075
1075
1075
1075
1075
1075
Динамические характеристики
175
183
173
182
183
182
183
182
182
12.6
10.7
13.4
11.6
10.7
11.6
10.7
11.6
11.6
17.0
15.2
10.2
8.8
15.2
8.8
15.2
8.8
8.8
Масса (5 мест)
1570
1590
1610
1620
1590
1620
1590
1620
1620
Расход топлива
50
50
50
50
50
50
50
50
50
Спецификация
H0S6K4617
H0S6D2617
H0S6K461F
H0S6D261F
H0S6D2617
H0S6D261F
H0S6D261F
H0S6D261F
H0S6D261F
Kia Rio X (Киа Рио Х) 2021 — 2022 – технические характеристики: двигатель, габариты, клиренс
Двигатель и трансмиссия
1. 4 MPI
4 MPI
1.6 MPI
1.4 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
1.6 MPI
100
123
100
123
123
123
123
123
123
72.0 х 84.0
77.0 х 85.4
72.0 х 84.0
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
77.0 х 85.4
передний
передний
передний
передний
передний
передний
передний
передний
передний
10.5
10.5
10.5
10.5
10.5
10.5
10.5
10.5
10.5
132
151
132
151
151
151
151
151
151
100 @ 6000
123 @ 6300
100 @ 6000
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
123 @ 6300
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
Бензин, АИ 92-95
132 @ 4000
151 @ 4850
132 @ 4000
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
151 @ 4850
1. 4
4
1.6
1.4
1.6
1.6
1.6
1.6
1.6
1.6
1368
1591
1368
1591
1591
1591
1591
1591
1591
4
4
4
4
4
4
4
4
4
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Евро-5
Механика (6MT)
Механика (6MT)
Автомат (6AT)
Автомат (6AT)
Механика (6MT)
Автомат (6AT)
Автомат (6AT)
Автомат (6AT)
Автомат (6AT)
передний
передний
передний
передний
передний
передний
передний
передний
передний
12,6
10,7
13,4
11,6
10,7
11,6
10,7
11,6
11,6
5,9
6,6
6,6
6,8
6,6
6,8
6,6
6,8
6,8
Сухое однодисковое
Сухое однодисковое
Гидравлическая муфта
Гидравлическая муфта
Сухое однодисковое
Гидравлическая муфта
Сухое однодисковое
Гидравлическая муфта
Гидравлическая муфта
1. 6~1.7
6~1.7
1.6~1.7
6.7
6.7
1.6~1.7
6.7
1.6~1.7
6.7
6.7
Выбросы CO2
175
202
200
207
202
207
202
207
207
117
126
126
130
126
130
126
130
130
140
154
154
158
154
158
154
158
158
Размеры
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
Хэтчбек
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
4275 / 1750 / 1535
2600
2600
2600
2600
2600
2600
2600
2600
2600
190
190
190
190
190
190
190
190
195
390
390
390
390
390
390
390
390
390
Электрооборудование
60
60
60
60
60
60
60
60
60
13. 5 В 90 А
5 В 90 А
13.5 В 120 А
13.5 В 90 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
13.5 В 120 А
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
12 В 0.9 кВт
3.5
3.6
3.5
3.6
3.6
3.6
3.6
3.6
3.6
Рулевое управление
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
Рулевой привод с электроусилителем
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
14.53:1
2.7
2.7
2.7
2.7
2. 7
7
2.7
2.7
2.7
2.7
5.2
5.2
5.2
5.2
5.2
5.2
5.2
5.2
5.2
Тормоза
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
256 x 22
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
262 x 10
Внутренние размеры
390
390
390
390
390
390
390
390
390
1075
1075
1075
1075
1075
1075
1075
1075
1075
Динамические характеристики
175
183
173
182
183
182
183
182
182
12.6
10.7
13.4
11.6
10.7
11.6
10.7
11.6
11.6
17.0
15.2
10.2
8.8
15.2
8.8
15.2
8.8
8.8
Масса (5 мест)
1570
1590
1610
1620
1590
1620
1590
1620
1620
Расход топлива
50
50
50
50
50
50
50
50
50
Спецификация
H0S6K4617
H0S6D2617
H0S6K461F
H0S6D261F
H0S6D2617
H0S6D261F
H0S6D261F
H0S6D261F
H0S6D261F
Технические характеристики двигателя Киа Рио и разгон до 100
Объем двигателя и разгон до 100 км/час автомобиля Киа Рио. Наведены основные поколения и комплектации модели, а так же возможные вариации в пределах одной генерации.
Наведены основные поколения и комплектации модели, а так же возможные вариации в пределах одной генерации.
Характеристики агрегата чаще всего решают динамические свойства машины. Как правило, чем больше крутящий момент, тем быстрей стартует автомобиль и тем больше его ускорение. По данным производителя, до 100 км/час Киа Рио может разогнаться от 8,7 до 16,1 секунды.
Характеристики двигателя Kia Rio 2020, 4 поколение, седан, рестайлинг, FB
Модель производится с 08.2020 по нынешнее время.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 100 л.с., бензин, МКПП, передний привод | 1368 | 132 | 12,2 |
| 1.6 л, 123 л.с., бензин, МКПП, передний привод | 1591 | 151 | 10,3 |
| 1.4 л, 100 л.с., бензин, АКПП, передний привод | 1368 | 132 | 12,9 |
1. 6 л, 123 л.с., бензин, АКПП, передний привод 6 л, 123 л.с., бензин, АКПП, передний привод | 1591 | 151 | 11,2 |
Характеристики двигателя Kia Rio 2016, 4 поколение, седан, FB
Модель производится с 11.2016 по нынешнее время. Читайте обзор Kia Rio 2017.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 100 л.с., бензин, МКПП, передний привод | 1368 | 132 | 12,2 |
| 1.6 л, 123 л.с., бензин, МКПП, передний привод | 1591 | 151 | 10,3 |
| 1.4 л, 100 л.с., бензин, АКПП, передний привод | 1368 | 132 | 12,9 |
| 1.6 л, 123 л.с., бензин, АКПП, передний привод | 1591 | 151 | 11,2 |
Характеристики двигателя Kia Rio 2015, 3 поколение, хэчбек, рестайлинг, QB
Модель производится с 06.2015 по 09.2017.
| Модификация | Объем двигателя, см. куб. куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.6 л, 123 л.с., бензин, МКПП, передний привод | 1591 | 155 | 10,3 |
| 1.4 л, 107 л.с., бензин, МКПП, передний привод | 1396 | 135 | 11,6 |
| 1.6 л, 123 л.с., бензин, АКПП, передний привод | 1591 | 155 | 11,2 |
| 1.4 л, 107 л.с., бензин, АКПП, передний привод | 1396 | 135 | 13,6 |
Характеристики двигателя Kia Rio 2015, 3 поколение, седан, рестайлинг, QB
Модель производилась с 04.2015 по 09.2017. Читайте обзор Kia Rio 2015.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 107 л.с., бензин, МКПП, передний привод | 1396 | 135 | 11,5 |
| 1.6 л, 123 л.с., бензин, МКПП, передний привод | 1591 | 155 | 10,3 |
1. 6 л, 123 л.с., бензин, АКПП, передний привод 6 л, 123 л.с., бензин, АКПП, передний привод | 1591 | 155 | 11,2 |
| 1.4 л, 107 л.с., бензин, АКПП, передний привод | 1396 | 135 | 13,5 |
Характеристики двигателя Kia Rio 2012, 3 поколение, хэчбек, QB
Модель производилась с 03.2012 по 05.2015.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.6 л, 123 л.с., бензин, МКПП, передний привод | 1591 | 155 | 10,3 |
| 1.4 л, 107 л.с., бензин, МКПП, передний привод | 1396 | 135 | 11,5 |
| 1.4 л, 107 л.с., бензин, АКПП, передний привод | 1396 | 135 | 13,5 |
| 1.6 л, 123 л.с., бензин, АКПП, передний привод | 1591 | 155 | 11,2 |
| 1.4 л, 90 л.с., дизель, МКПП, передний привод | 1396 | 220 | 14,2 |
| 1. 1 л, 75 л.с., дизель, МКПП, передний привод | 1120 | 170 | 16,1 |
| 1.2 л, 86 л.с., бензин, МКПП, передний привод | 1248 | 121 | 13,1 |
| 1.4 л, 109 л.с., бензин, МКПП, передний привод | 1396 | 137 | 11,5 |
| 1.4 л, 109 л.с., бензин, АКПП, передний привод | 1396 | 137 | 13,2 |
Характеристики двигателя Kia Rio 2011, 3 поколение, 3 двери, хэчбек, UB
Модель производилась с 03.2011 по 05.2015.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 90 л.с., дизель, МКПП, передний привод | 1396 | 220 | 14,2 |
| 1.1 л, 75 л.с., дизель, МКПП, передний привод | 1120 | 170 | 16,1 |
| 1.2 л, 86 л.с., бензин, МКПП, передний привод | 1248 | 121 | 13,1 |
1. 4 л, 109 л.с., бензин, МКПП, передний привод 4 л, 109 л.с., бензин, МКПП, передний привод | 1396 | 137 | 11,5 |
| 1.4 л, 109 л.с., бензин, АКПП, передний привод | 1396 | 137 | 13,2 |
Характеристики двигателя Kia Rio 2011, 3 поколение, седан, QB
Модель производилась с 08.2011 по 03.2015.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 107 л.с., бензин, МКПП, передний привод | 1396 | 135 | 11,5 |
| 1.6 л, 123 л.с., бензин, МКПП, передний привод | 1591 | 155 | 10,3 |
| 1.4 л, 107 л.с., бензин, АКПП, передний привод | 1396 | 135 | 13,5 |
| 1.6 л, 123 л.с., бензин, АКПП, передний привод | 1591 | 155 | 11,2 |
Характеристики двигателя Kia Rio 2011 для США, 3 поколение, хэчбек, UB
Модель производилась с 03. 2011 по 05.2015.
2011 по 05.2015.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.6 л, 138 л.с., бензин, АКПП, передний привод | 1591 | 167 | 9,7 |
| 1.6 л, 138 л.с., бензин, МКПП, передний привод | 1591 | 167 | 8,7 |
Характеристики двигателя Kia Rio 2011 для США, 3 поколение, седан, UB
Модель производилась с 03.2011 по 03.2015.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.6 л, 138 л.с., бензин, АКПП, передний привод | 1591 | 167 | 9,3 |
| 1.6 л, 138 л.с., бензин, МКПП, передний привод | 1591 | 167 | 8,8 |
Характеристики двигателя Kia Rio 2009, 2 поколение, хэчбек, рестайлинг, JB
Модель производилась с 09. 2009 по 08.2011.
2009 по 08.2011.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 95 л.с., бензин, МКПП, передний привод | 1399 | 125 | 12,3 |
| 1.4 л, 95 л.с., бензин, АКПП, передний привод | 1399 | 125 | 14,5 |
| 1.5 л, 110 л.с., дизель, МКПП, передний привод | 1493 | 235 | 11,5 |
| 1.4 л, 97 л.с., бензин, МКПП, передний привод | 1399 | 125 | 12,3 |
| 1.6 л, 112 л.с., бензин, МКПП, передний привод | 1599 | 146 | 10,2 |
| 1.4 л, 75 л.с., бензин, МКПП, передний привод | 1399 | 125 | 12,3 |
| 1.6 л, 112 л.с., бензин, АКПП, передний привод | 1599 | 146 | 12 |
Характеристики двигателя Kia Rio 2009, 2 поколение, седан, рестайлинг, JB
Модель производилась с 09. 2009 по 08.2011.
2009 по 08.2011.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 95 л.с., бензин, МКПП, передний привод | 1399 | 125 | 12,3 |
| 1.4 л, 95 л.с., бензин, АКПП, передний привод | 1399 | 125 | 14,5 |
Характеристики двигателя Kia Rio 2005, 2 поколение, хэчбек, JB
Модель производилась с 03.2005 по 08.2009.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 97 л.с., бензин, МКПП, передний привод | 1399 | 125 | 12,3 |
| 1.4 л, 97 л.с., бензин, АКПП, передний привод | 1399 | 125 | 14,5 |
| 1.5 л, 110 л.с., дизель, МКПП, передний привод | 1493 | 235 | 11,5 |
1. 6 л, 112 л.с., бензин, МКПП, передний привод 6 л, 112 л.с., бензин, МКПП, передний привод | 1599 | 146 | 10,2 |
| 1.6 л, 112 л.с., бензин, АКПП, передний привод | 1599 | 146 | 10,6 |
Характеристики двигателя Kia Rio 2005, 2 поколение, седан, JB
Модель производилась с 03.2005 по 08.2009.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.4 л, 97 л.с., бензин, МКПП, передний привод | 1399 | 125 | 12,3 |
| 1.4 л, 97 л.с., бензин, АКПП, передний привод | 1399 | 125 | 14,5 |
Характеристики двигателя Kia Rio 2009 для США, 2 поколение, хэчбек, рестайлинг, JB
Модель производилась с 09.2009 по 08.2011.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
1. 6 л, 110 л.с., бензин, АКПП, передний привод 6 л, 110 л.с., бензин, АКПП, передний привод | 1599 | 145 | — |
| 1.6 л, 110 л.с., бензин, МКПП, передний привод | 1599 | 145 | — |
Характеристики двигателя Kia Rio 2009 для США, 2 поколение, седан, рестайлинг, JB
Модель производилась с 09.2009 по 08.2011.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.6 л, 110 л.с., бензин, АКПП, передний привод | 1599 | 145 | — |
| 1.6 л, 110 л.с., бензин, МКПП, передний привод | 1599 | 145 | — |
Характеристики двигателя Kia Rio 2005 для США, 2 поколение, хэчбек, JB
Модель производилась с 03.2005 по 08.2009.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
1. 6 л, 110 л.с., бензин, АКПП, передний привод 6 л, 110 л.с., бензин, АКПП, передний привод | 1599 | 145 | — |
| 1.6 л, 110 л.с., бензин, МКПП, передний привод | 1599 | 145 | — |
Характеристики двигателя Kia Rio 2005 для США, 2 поколение, седан, JB
Модель производилась с 03.2005 по 08.2009.
| Модификация | Объем двигателя, см.куб. | Максимальный крутящий момент, Н*м | Разгон до 100 км/ч, с |
| 1.6 л, 110 л.с., бензин, АКПП, передний привод | 1599 | 145 | — |
| 1.6 л, 110 л.с., бензин, МКПП, передний привод | 1599 | 145 | — |
Объем двигателя и разгон до 100 км/час автомобиля Киа Рио. Наведены основные поколения и комплектации модели, а так же возможные вариации в пределах одной генерации.
||list|
Посмотреть другие характеристики Kia Rio:
Генерации Kia Rio:
- 4 поколение 2020 (FB, седан, рестайлинг)
- 4 поколение 2016 (FB, седан)
- 3 поколение 2015-2017 (QB, хэтчбек, ресталийнг)
- 3 поколение 2015-2017 (QB, седан, ресталийнг)
- 3 поколение 2012-2015 (QB, хэтчбек)
- 3 поколение 2011-2015 (UB, хэтчбек, 3DR)
- 3 поколение 2011-2015 (QB, седан)
- 3 поколение 2011-2015 (UB, хэтчбек, для США)
- 3 поколение 2011-2015 (UB, седан, для США)
- 2 поколение 2009-2011 (JB, хэтчбек, ресталийнг)
- 2 поколение 2009-2011 (JB, седан, ресталийнг)
- 2 поколение 2005-2009 (JB, хэтчбек)
- 2 поколение 2005-2009 (JB, седан)
- 2 поколение 2009-2011 (JB, хэтчбек, рестайлинг, для США)
- 2 поколение 2009-2011 (JB, седан, рестайлинг, для США)
- 2 поколение 2005-2009 (JB, хэтчбек, для США)
- 2 поколение 2005-2009 (JB, седан, для США)
Kia
Сигнальная реактивность связана с продолжительностью и тяжестью алкогольной зависимости: исследование фМРТ
1. Лешнер А.И. (1997) Наркомания – это болезнь мозга, и это имеет значение. Наука
278: 45–47. [PubMed] [Google Scholar]
Лешнер А.И. (1997) Наркомания – это болезнь мозга, и это имеет значение. Наука
278: 45–47. [PubMed] [Google Scholar]
2. Маклеллан А.Т., Льюис Д.К., О’Брайен С.П., Клебер Х.Д. (2000)Зависимость от наркотиков, хроническое заболевание: последствия для лечения, страхования и оценки результатов. ДЖАМА 284: 1689–1695. [PubMed] [Google Scholar]
3. Американская психиатрическая ассоциация (2001 г.) Диагностическое и статистическое руководство по психическим расстройствам, четвертое издание. Вашингтон, округ Колумбия: Американское психиатрическое издательство.
4. Heilig M, Thorsell A, Sommer WH, Hansson AC, Ramchandani VA, et al. (2010) Превращение нейробиологии алкоголизма в клиническое лечение: от блокирования гудения до лечения хандры. Neurosci Biobehav Rev 35: 334–344. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5.
Эверитт Б.Дж., Роббинс Т.В. (2005)Нейронные системы подкрепления при наркомании: от действий к привычкам к принуждению. Нат Нейроски
8: 1481–1489 nn1579 [pii]; 10. 1038/nn1579 [doi]. [PubMed] [Академия Google]
1038/nn1579 [doi]. [PubMed] [Академия Google]
6. Робинсон Т.Е., Берридж К.С. (1993) Нейронная основа тяги к наркотикам: теория зависимости от стимулов и сенсибилизации. Мозг Res Мозг Res Rev 18: 247–291. [PubMed] [Google Scholar]
7. Монти П.М., Бинкофф Дж.А., Абрамс Д.Б., Цвик В.Р., Ниренберг Т.Д. и др. (1987) Реакция алкоголиков и неалкоголиков на признаки употребления алкоголя. J Abnorm Psychol 96: 122–126. [PubMed] [Google Scholar]
8. Дикинсон А., Смит Дж., Миренович Дж. (2000) Диссоциация павловского и инструментального поощрительного обучения под действием антагонистов допамина. Поведение Нейроски 114: 468–483. [PubMed] [Академия Google]
9. Тиффани С.Т., Картер Б.Л. (1998) Является ли тяга источником компульсивного употребления наркотиков? Журнал психофармакологии 12: 23–30. [PubMed] [Google Scholar]
10.
Хайнц А., Бек А., Груссер С.М., Грейс А.А., Враз Дж. (2009) Выявление нейронной схемы тяги к алкоголю и уязвимости к рецидивам. Наркоман Биол
14: 108–118. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Демос К.Е., Хизертон Т.Ф., Келли В.М. (2012) Индивидуальные различия в активности прилежащего ядра в отношении еды и сексуальных изображений предсказывают увеличение веса и сексуальное поведение. Джей Нейроски 32: 5549–5552. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Murdaugh DL, Cox JE, Cook EW, Weller RE (2012) МРТ-реактивность изображений высококалорийной пищи предсказывает краткосрочные и долгосрочные результаты программы по снижению веса. Нейроизображение 59: 2709–2721. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Ito R, Dalley JW, Robbins TW, Everitt BJ (2002) Выброс дофамина в дорсальном стриатуме во время поведения, связанного с поиском кокаина, под контролем сигнала, связанного с наркотиками. . J Neurosci 22: 6247–6253. 20026606 [doi]; 14.22.6247 [pii]. [Бесплатная статья PMC] [PubMed]
14.
Vanderschuren LJMJ, Di Ciano P, Everitt BJ (2005) Участие дорсального полосатого тела в поиске кокаина, контролируемом сигналом. Джей Нейроски
25: 8665–8670. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Джей Нейроски
25: 8665–8670. [Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Гараван Х., Панкевич Дж., Блум А., Чо Дж.К., Сперри Л. и др. (2000) Тяга к кокаину, вызванная сигналом: нейроанатомическая специфичность для потребителей наркотиков и стимулы для наркотиков. Эм Джей Психиатрия 157: 1789–1798. [PubMed] [Google Scholar]
16. Волков Н.Д., Ван Г.Дж., Теланг Ф., Фаулер Дж.С., Логан Дж. и др. (2006) Кокаиновые сигналы и дофамин в дорсальном полосатом теле: механизм тяги при кокаиновой зависимости. Джей Нейроски 26: 6583–6588 24/26/6583 [pii]; 10.1523/JNEUROSCI.1544-06.2006 [doi]. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Волков Н.Д., Ван Г.Дж., Теланг Ф., Фаулер Дж.С., Логан Дж. и др. (2008) Повышение уровня дофамина в полосатом теле не вызывает тягу у лиц, злоупотребляющих кокаином, если только они не сопровождаются кокаиновыми сигналами. Нейроизображение 39: 1266–1273 S1053-8119(07)00881-6 [pii];10.1016/j.neuroimage.2007.09.059 [doi]. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Вонг Д.Ф., Кувабара Х., Шретлен Д.Дж., Бонсон К.Р., Чжоу Ю и др. (2006) Повышенная занятость дофаминовых рецепторов в полосатом теле человека во время вызванной сигналом тяги к кокаину. нейропсихофармакология
31: 2716–2727 1301194 [pii];10.1038/sj.npp.1301194 [doi]. [PubMed] [Google Scholar]
Вонг Д.Ф., Кувабара Х., Шретлен Д.Дж., Бонсон К.Р., Чжоу Ю и др. (2006) Повышенная занятость дофаминовых рецепторов в полосатом теле человека во время вызванной сигналом тяги к кокаину. нейропсихофармакология
31: 2716–2727 1301194 [pii];10.1038/sj.npp.1301194 [doi]. [PubMed] [Google Scholar]
19. Фоллштадт-Кляйн С., Вихерт С., Рабинштейн Дж., Бюлер М., Кляйн О. и соавт. (2010) Первоначальное, привычное и компульсивное употребление алкоголя характеризуется смещением обработки сигналов с вентрального на дорсальный полосатый телес. Зависимость 105: 1741–1749. [PubMed] [Google Scholar]
20. Schacht J, Anton R, Myrick H (2012) Функциональные нейровизуализационные исследования реактивности алкогольных сигналов: количественный метаанализ и систематический обзор. Наркоман биол. [Бесплатная статья PMC] [PubMed]
21.
Boschloo L, Vogelzangs N, Smit JH, van den Brink W, Veltman DJ и др. (2011) Показатели сопутствующих заболеваний и риска расстройств, связанных с употреблением алкоголя, среди лиц с тревожными и/или депрессивными расстройствами. Результаты Нидерландского исследования депрессии и тревоги (NESDA). J влияет на расстройство
131: 233–242 S0165-0327(10)00749-4 [pii];10.1016/j.jad.2010.12.014 [doi]. [PubMed] [Google Scholar]
Результаты Нидерландского исследования депрессии и тревоги (NESDA). J влияет на расстройство
131: 233–242 S0165-0327(10)00749-4 [pii];10.1016/j.jad.2010.12.014 [doi]. [PubMed] [Google Scholar]
22. Пиццагалли Д.А., Иосифеску Д., Халлетт Л.А., Ратнер К.Г., Фава М. (2008)Сниженная гедонистическая способность при большом депрессивном расстройстве: данные вероятностной задачи вознаграждения. J Psychiatr Res 43: 76–87. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Эшель Н., Ройзер Дж. П. (2010) Поощрение и наказание при депрессии. биопсихиатрия 68: 118–124. [PubMed] [Google Scholar]
24. Penninx BWJH, Beekman ATF, Smit JH, Zitman FG, Nolen WA и др. (2008) Нидерландское исследование депрессии и тревоги (NESDA): обоснование, цели и методы. Int J Methods Psychiatr Res 17: 121–140. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25.
Робинс Л.Н., Винг Дж., Виттхен Х.У., Хельцер Дж.Е., Бабор Т.Ф. и соавт. (1988) Составное международное диагностическое интервью. Эпидемиологический инструмент, пригодный для использования в сочетании с различными диагностическими системами и в разных культурах. Arch Gen Психиатрия
45: 1069–1077. [PubMed] [Google Scholar]
Эпидемиологический инструмент, пригодный для использования в сочетании с различными диагностическими системами и в разных культурах. Arch Gen Психиатрия
45: 1069–1077. [PubMed] [Google Scholar]
26. Sullivan JT, Sykora K, Schneiderman J, Naranjo CA, Sellers EM (1989)Оценка синдрома отмены алкоголя: пересмотренная оценка клинического института по алкогольной шкале (CIWA-Ar). бр Джей наркоман 84: 1353–1357. [PubMed] [Google Scholar]
27. Van Strien JW (1992) Classificatie van links-en rechthandige proefpersonen [Классификация левшей и правшей участников исследования]. Нед Тиджшр Психол 47: 88–92. [Академия Google]
28. Babor TF, Kranzler HR, Lauerman RJ (1989) Раннее выявление вредного употребления алкоголя: сравнение клинических, лабораторных и самоотчетных процедур скрининга. Поведение наркомана 14: 139–157. [PubMed] [Google Scholar]
29.
Heatherton TF, Kozlowski LT, Frecker RC, Fagerstrom KO (1991) Тест Fagerstrom на зависимость от никотина: пересмотренная версия вопросника Fagerstrom Tolerance Questionnaire. бр Джей наркоман
86: 1119–1127. [PubMed] [Google Scholar]
бр Джей наркоман
86: 1119–1127. [PubMed] [Google Scholar]
30. Бек А.Т., Эпштейн Н., Браун Г., Стир Р.А. (1988) Инвентарь для измерения клинической тревожности: психометрические свойства. J Consult Clin Psychol 56: 893–897. [PubMed] [Google Scholar]
31. Rush AJ, Gullion CM, Basco MR, Jarrett RB, Trivedi MH (1996) Перечень депрессивной симптоматики (IDS): психометрические свойства. Психол Мед 26: 477–486. [PubMed] [Google Scholar]
32. Лав А., Джеймс Д., Уилнер П. (1998) Сравнение двух анкет о тяге к алкоголю. Зависимость 93: 1091–1102. [PubMed] [Академия Google]
33. Дейхманн Р., Готфрид Дж. А., Хаттон С., Тернер Р. (2003) Оптимизированный EPI для фМРТ-исследований орбитофронтальной коры. Нейроизображение 19: 430–441. [PubMed] [Google Scholar]
34. Фристон К.Дж., Холмс А.П., Полайн Дж.Б., Грасби П.Дж., Уильямс С.К. и др. (1995) Повторный анализ временных рядов фМРТ. Нейроизображение 2: 45–53. [PubMed] [Google Scholar]
35.
Zijlstra F, Veltman DJ, Booij J, van den Brink W, Franken IHA (2009)Нейробиологические субстраты влечения, вызванного сигналом, и ангедонии у недавно воздерживающихся от опиоидов мужчин. Наркотики Алкоголь Зависит
99: 183–192. [PubMed] [Google Scholar]
Наркотики Алкоголь Зависит
99: 183–192. [PubMed] [Google Scholar]
36. Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, et al. (2002)Автоматизированная анатомическая маркировка активаций в SPM с использованием макроскопической анатомической парцелляции мозга одного субъекта MNI MRI. Нейроизображение 15: 273–289 10.1006/nimg.2001.0978 [doi]; S1053811
9784 [pii]. [PubMed] [Google Scholar]37. Maldjian JA, Laurienti PJ, Kraft RA, Burdette JH (2003) Автоматизированный метод нейроанатомического и цитоархитектонического анализа наборов данных фМРТ на основе атласа. Нейроизображение 19: 1233–1239. S1053811
1691 [pii]. [PubMed]
38. Nielsen FA, Hansen LK (2002) Автоматическая анатомическая маркировка координат Talairach и создание интересующих объемов с помощью базы данных BrainMap (представлено на 8-й Международной конференции по функциональному картированию человеческого мозга, 2–6 июня). , 2002, Сендай, Япония (есть на компакт-диске). Нейроимидж 16.:no_upscale()/imgs/2015/04/09/10/132410/0e79ff0455b175ccea32942b2596b27c328ac8c9.jpg)
39. Fox PT, Lancaster JL (1994) Неврология в сети. Наука 266: 994–996. [PubMed] [Google Scholar]
40. де Вит С., Уотсон П., Харсей Х.А., Коэн М.Х., ван де Вийвер И. и др. (2012) Кортико-стриарная связь лежит в основе индивидуальных различий в балансе между привычным и целенаправленным контролем действий. Джей Нейроски 32: 12066–12075. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Tricomi E, Balleine BW, O’Doherty JP (2009)Особая роль заднего дорсолатерального полосатого тела в обучении привычкам человека. Eur J Neurosci 29: 2225–2232 EJN6796 [pii];10.1111/j.1460-9568.2009.06796.x [doi]. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Уорсли К.Дж., Марретт С., Нилин П., Вандал А.С., Фристон К.Дж. и др. (1996) Единый статистический подход для определения значимых сигналов на изображениях церебральной активации. Hum Brain Map 4: 58–73. [PubMed] [Академия Google]
43.
Lieberman MD, Cunningham WA (2009) Проблемы с ошибками типа I и типа II в исследованиях фМРТ: перебалансировка шкалы. Soc Cogn влияет на нейроны
4: 423–428 nsp052 [pii]; 10.1093/scan/nsp052 [doi]. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Soc Cogn влияет на нейроны
4: 423–428 nsp052 [pii]; 10.1093/scan/nsp052 [doi]. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Груссер С.М., Враз Дж., Кляйн С., Герман Д., Смолка М.Н. и соавт. (2004) Индуцированная сигналом активация полосатого тела и медиальной префронтальной коры связана с последующим рецидивом у трезвенников. Психофармакология (Берл) 175: 296–302 10.1007/s00213-004-1828-4 [doi]. [PubMed] [Академия Google]
45. Хайнц А., Зиссмайер Т., Враз Дж., Герман Д., Кляйн С. и соавт. (2004) Корреляция между дофаминовыми рецепторами D(2) в вентральном полосатом теле и центральной обработкой алкогольных сигналов и тяги. Эм Джей Психиатрия 161: 1783–1789 161/10/1783 [pii]; 10.1176/appi.ajp.161.10.1783 [doi]. [PubMed] [Google Scholar]
46. Braus DF, Wrase J, Grusser S, Hermann D, Ruf M, et al. (2001)Ассоциированные алкоголем стимулы активируют вентральное полосатое тело у трезвенников. J нейронная передача 108: 887–894. [PubMed] [Google Scholar]
47. Fontenelle LF, Oostermeijer S, Harrison BJ, Pantelis C, Yucel M (2011)Обсессивно-компульсивное расстройство, расстройства импульсного контроля и наркомания: общие черты и возможные методы лечения. Наркотики
71: 827–840. [PubMed] [Google Scholar]
Fontenelle LF, Oostermeijer S, Harrison BJ, Pantelis C, Yucel M (2011)Обсессивно-компульсивное расстройство, расстройства импульсного контроля и наркомания: общие черты и возможные методы лечения. Наркотики
71: 827–840. [PubMed] [Google Scholar]
48. Sjoerds Z, de Wit S, van den Brink W, Robbins TW, Beekman AT и др.. (2013) Поведенческие и нейровизуализационные данные о чрезмерной зависимости от обучения привычкам у пациентов с алкогольной зависимостью. Трансляционная психиатрия в прессе. [Бесплатная статья PMC] [PubMed]
49. Сайетт М.А., Шиффман С., Тиффани С.Т., Ниаура Р.С., Мартин С.С. и др. (2000) Измерение тяги к наркотикам. Зависимость 95 Приложение 2С189–С210. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50.
Фельдштейн Юинг С.В., Филби Ф.М., Чандлер Л.Д., Хатчисон К.Е. (2010)Изучение взаимосвязи между симптомами депрессии и тревоги и реакцией нейронов на алкогольные сигналы. Алкоголь Clin Exp Res
34: 396–403 ACER1104 [pii];10.1111/j.1530-0277.2009.01104. x [doi]. [Бесплатная статья PMC] [PubMed] [Google Scholar]
x [doi]. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Breese GR, Sinha R, Heilig M (2011) Хроническая алкогольная нейроадаптация и стресс способствуют восприимчивости к тяге к алкоголю и рецидивам. Фармакол Тер 129: 149–171 S0163-7258(10)00191-9 [pii];10.1016/j.pharmthera.2010.09.007 [doi]. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Выбор подсказки как функция нехватки времени и предполагаемого риска
РЕЗЮМЕ. Сигналы используются людьми при формировании убеждений о брендах, которые, в свою очередь, влияют на выбор бренда. Чтобы предсказать и понять, почему люди демонстрируют определенное поведение при выборе бренда, необходимо понять, почему люди используют определенные типы сигналов. В этой статье описывается исследование, в котором изучалось влияние нехватки времени и предполагаемого риска на выбор людей между двумя типами сигналов (сигналы с высокой прогностической ценностью, но с низкой достоверностью, и сигналы с низкой прогностической ценностью, но с высокой достоверностью). Эксперимент имел значительный основной эффект для нехватки времени и значительный срок взаимодействия.
Эксперимент имел значительный основной эффект для нехватки времени и значительный срок взаимодействия.
Ссылка:
Дуглас А. (Тони) Шеллинк (1983), «Выбор сигнала как функция временного давления и предполагаемого риска», в NA — Advances in Consumer Research Volume 10, eds. Ричард П. Багоцци и Элис М. Тайбут, Анн-Абор, Мичиган: Ассоциация потребительских исследований, страницы: 470-475.
Достижения в исследованиях потребителей, том 10, 1983 г. Страницы 470–475
ВЫБОР РЕКОМЕНДИЙ КАК ФУНКЦИЯ ОТ ВРЕМЕНИ И ВОСПРИНИМАЕМОГО РИСКА
Дуглас А. (Тони) Шеллинк, Университет Далхаузи
[Автор благодарит Джагдиша Шета, Дэвида Гарднера и Рассела Белка за их руководство в проведении исследования, описанного в этой статье .]
ABSTRACT —
Сигналы используются людьми при формировании убеждений о брендах, которые, в свою очередь, влияют на выбор бренда. Чтобы предсказать и понять, почему люди демонстрируют определенное поведение при выборе бренда, необходимо понять, почему люди используют определенные типы сигналов. В этой статье описывается исследование, в котором изучалось влияние нехватки времени и предполагаемого риска на выбор людей между двумя типами сигналов (сигналы с высокой прогностической ценностью, но с низкой достоверностью, и сигналы с низкой прогностической ценностью, но с высокой достоверностью). Эксперимент имел значительный основной эффект для нехватки времени и значительный срок взаимодействия.
Чтобы предсказать и понять, почему люди демонстрируют определенное поведение при выборе бренда, необходимо понять, почему люди используют определенные типы сигналов. В этой статье описывается исследование, в котором изучалось влияние нехватки времени и предполагаемого риска на выбор людей между двумя типами сигналов (сигналы с высокой прогностической ценностью, но с низкой достоверностью, и сигналы с низкой прогностической ценностью, но с высокой достоверностью). Эксперимент имел значительный основной эффект для нехватки времени и значительный срок взаимодействия.
ВВЕДЕНИЕ
На сегодняшний день было проведено мало исследований детерминант поведения при выборе сигнала. Было проведено значительное исследование того, какие сигналы люди обычно выбирают, но не почему они их выбирают (Bettman and Jacoby 1975, Jacoby, Chestnut, Weisl and Fisher 1975, Jacoby, Szybillo and Bersato-Schach 1977, Kohn and Jacoby 1973, Olson and Джейкоби 1972, Пинкус и Уотерс 1975). Из этого исследования становятся ясными две точки дождя. Во-первых, при вынесении суждения люди обычно обращают внимание на меньшее, чем полное количество сигналов, которые им доступны. Во-вторых, при оценке качества бренда они склонны учитывать название бренда и внутренние признаки продукта (сигналы, которые нельзя изменить без изменения объекта).
Из этого исследования становятся ясными две точки дождя. Во-первых, при вынесении суждения люди обычно обращают внимание на меньшее, чем полное количество сигналов, которые им доступны. Во-вторых, при оценке качества бренда они склонны учитывать название бренда и внутренние признаки продукта (сигналы, которые нельзя изменить без изменения объекта).
Это поднимает вопрос о том, как потребитель делает выбор из доступного набора сигналов и почему он часто полагается на сигналы, которые, по их собственному признанию, скорее всего не предсказывают характеристики рассматриваемых брендов? Целью данного исследования является проверка влияния двух возможных детерминант поведения при выборе сигнала.
Сигнал определяется как характеристика, событие, качество или объект, внешние по отношению к человеку, которые могут быть закодированы и использованы для классификации объекта-стимула (Schellinck 1980). Существуют сотни потенциальных сигналов, которые можно связать с любым конкретным объектом. Что требуется, так это какие-то средства категоризации или описания сигналов, чтобы можно было изучить детерминанты выбора среди этих различных типов сигналов.
Что требуется, так это какие-то средства категоризации или описания сигналов, чтобы можно было изучить детерминанты выбора среди этих различных типов сигналов.
При обсуждении сигналов используется несколько таксономий, большинство из которых похожи, но получены из разных областей исследования. К ним относятся внутреннее/внешнее (Эндрюс и Валенци, 1971; Олсон и Джейкоби, 1972), фигура/фон (например, Тверски и Канеман, 19).80), описательные/поведенческие (Сарбин, Тафт и Бейли, 1960) и сигнифицирующие/символические (Ховард и Шет, 1969).
Вместо того, чтобы классифицировать сигналы, их можно охарактеризовать. Опять же, ярлыки пришли из разных областей исследований, некоторые из которых по существу описывают одни и те же характеристики. Дескрипторы в исследовании сигналов включают очевидность и актуальность сигнала (Archer, 1962), избыточность сигнала (Rollings, Bethel and Deffenbacker, 1971, Einhorn, Kleinmertz, and Kleinmertz, 1979), общность сигнала (Slovic and MacPhillany, 19). 74) экологическая достоверность сигнала (Брансуик, 1949; Винак и Аркофф, 1957; Келли и Арровуд, 1960); обобщаемость сигналов (Брим, 1954; Френч и Снайдер, 1959); достоверность суждений и достоверность сигнала (Брунер, Гудноу и др.). Остин, 1956), а также достоверность и прогностическая ценность сигналов (Кокс, 1967, Олсон и Джейкоби, 1972).
74) экологическая достоверность сигнала (Брансуик, 1949; Винак и Аркофф, 1957; Келли и Арровуд, 1960); обобщаемость сигналов (Брим, 1954; Френч и Снайдер, 1959); достоверность суждений и достоверность сигнала (Брунер, Гудноу и др.). Остин, 1956), а также достоверность и прогностическая ценность сигналов (Кокс, 1967, Олсон и Джейкоби, 1972).
Можно было бы использовать многие из этих схем категоризации, но правило сортировки сигналов, разработанное Коксом (1967) для изучения поведения покупателей, использует прогностическую и доверительную ценность. Последующие исследования Олсона и Джейкоби (1972) показал, что эта схема характеристики сигналов полезна для прогнозирования того, какие сигналы люди будут использовать при оценке брендов.
Эта схема характеристики сигналов особенно подходит для изучения поведения покупателей с точки зрения потребительства. Защитники прав потребителей, как правило, настаивают на предоставлении покупателям сигналов, обладающих высокой прогностической ценностью, при условии, что потребители будут использовать эти сигналы. Однако потребители часто игнорируют эти сигналы. Если можно показать, что на выбор потребителем сигналов влияет значение достоверности сигнала при некоторых обстоятельствах, то потребители и правительственные учреждения, которые регулируют требования к информации для потребителей, могут принять этот факт во внимание. В это «иррациональное» поведение можно пролить свет.
Однако потребители часто игнорируют эти сигналы. Если можно показать, что на выбор потребителем сигналов влияет значение достоверности сигнала при некоторых обстоятельствах, то потребители и правительственные учреждения, которые регулируют требования к информации для потребителей, могут принять этот факт во внимание. В это «иррациональное» поведение можно пролить свет.
В опубликованном здесь исследовании изучалось использование подсказок с разным уровнем достоверности и прогностической ценности. Кокс (1967) определил прогностическую ценность как воспринимаемую вероятность, с которой категория критерия связана с категорией атрибута, а значение достоверности как воспринимаемую вероятность, с которой каждый сигнал может быть правильно отсортирован индивидуумом в категорию критерия.
Упомянутая ранее работа по использованию подсказок показывает, что люди используют такие подсказки, как торговая марка, страна производства или цена, даже когда потребитель знает, что они не особенно предсказывают качество. Кокс определил другие признаки (запах чулок, указывающий на качество носки; цвет коробок с моющим средством для одежды, указывающий на чистящую способность; громкость электродвигателей, указывающий на их мощность; и цвет растительного масла, указывающий на его густоту). люди используют, но которые имеют низкую прогностическую ценность. Он чувствует, что эти сигналы используются, хотя их прогностическая ценность невелика, потому что они обладают высокой достоверностью по сравнению с другими доступными сигналами.
Кокс определил другие признаки (запах чулок, указывающий на качество носки; цвет коробок с моющим средством для одежды, указывающий на чистящую способность; громкость электродвигателей, указывающий на их мощность; и цвет растительного масла, указывающий на его густоту). люди используют, но которые имеют низкую прогностическую ценность. Он чувствует, что эти сигналы используются, хотя их прогностическая ценность невелика, потому что они обладают высокой достоверностью по сравнению с другими доступными сигналами.
Олсон и Джейкоби (1972) показали, что потребители в первую очередь будут выбирать сигналы с высокой прогностической ценностью и высокой достоверностью, когда они легко доступны. После того, как эти сигналы закодированы, а у потребителя все еще слишком много неопределенности, чтобы принять решение, он должен выбрать сигналы с более высокой прогностической ценностью и более низким значением достоверности или сигналы с более низкой прогностической ценностью и более высоким значением достоверности. Вопрос в том, какие факторы заставят их предпочесть один тип сигнала другому.
Вопрос в том, какие факторы заставят их предпочесть один тип сигнала другому.
Выбор сигнала, который силен в одном измерении и слаб в другом, имеет свой тип риска, связанный с каждым типом сигнала. При наличии сигналов с высокой прогностической ценностью/низкой достоверностью существует более высокий риск того, что лицо, принимающее решения, полностью неверно интерпретирует данные и, таким образом, сделает неправильный выбор. Однако, если данные интерпретированы правильно, шансы на выбор отличного бренда выше при использовании сигналов с высокой достоверностью/низкой прогностической ценностью. Таким образом, использование сигналов с высокой прогностической ценностью и низкой достоверностью связано с более высоким риском, но потенциально с большей отдачей.
Выбор подсказки с высокой достоверностью/низкой прогностической ценностью означает, что меньше шансов неправильно определить, что, согласно подсказкам, должно быть лучшим брендом. Но шансов, что реплики указывают на лучшую марку, меньше. Таким образом, риск неправильной интерпретации сигналов меньше, но и потенциальная отдача меньше.
Таким образом, риск неправильной интерпретации сигналов меньше, но и потенциальная отдача меньше.
Предполагается, что когда люди сильно мотивированы на выбор хорошего бренда, они будут склонны выбирать сигналы с высокой прогностической ценностью/низкой достоверностью, а не с высокой достоверностью/низкой прогностической ценностью. Покупатели заинтересованы в том, чтобы приложить усилия, необходимые для тщательного анализа и интерпретации сигналов. Они чувствуют необходимость использовать подсказки, которые максимизируют их шансы сделать правильный выбор. Если люди не заинтересованы в выборе отличного бренда или сталкиваются с ограничениями, которые ограничивают их способность анализировать сигналы, они склонны приобретать ценность доверия; то есть они будут склонны выбирать сигналы с высокой достоверностью/низкой прогностической ценностью вместо сигналов с высокой прогностической ценностью/низкой достоверностью.
Целью данного исследования было проверить эти общие гипотезы путем изучения влияния мотиватора и ограничения на выбор сигналов покупателем. Мотивирующим фактором был предполагаемый риск, а ограничением – давление на зубья. В частности, было выдвинуто предположение, что:
Мотивирующим фактором был предполагаемый риск, а ограничением – давление на зубья. В частности, было выдвинуто предположение, что:
1. По мере увеличения нехватки времени будет возрастать тенденция зависимости от сигналов с высокой достоверностью/низкой прогностической ценностью.
Было высказано предположение, что, учитывая нехватку времени, люди будут чувствовать, что у них нет времени на тщательный анализ сигналов с высокой прогностической ценностью и низкой достоверностью. Они могут видеть ценность использования сигналов с высокой прогностической ценностью и низкой достоверностью, но чувствуют, что риск неправильной интерпретации сигналов слишком высок, и у них нет времени, чтобы тщательно проанализировать сигналы и снизить этот риск. Использование сигналов с высокой достоверностью/низкой прогностической ценностью, вероятно, считается более эффективным с точки зрения времени. Хотя шансы сделать лучший выбор уменьшаются, по крайней мере, они с большей вероятностью сделают удовлетворительный выбор.
2. По мере увеличения воспринимаемого риска существует тенденция полагаться на сигналы с высокой прогностической ценностью/низкой достоверностью.
Предполагается, что по мере увеличения воспринимаемой изменчивости в эффективности бренда покупатели чувствуют, что они должны больше полагаться на сигналы с высокой прогностической ценностью/низкой достоверностью, чтобы идентифицировать лучшие бренды. Это означает, что, поскольку производительность сильно различается, они с большей вероятностью смогут обнаружить существенные различия в значениях сигналов и, следовательно, определить, какой бренд является хорошим, а какой — плохим. Таким образом, шансы неверно истолковать сигналы кажутся уменьшенными.
Кроме того, при высокой неопределенности производительности увеличиваются шансы выбрать неудовлетворительную марку. Если цена принятия неправильного решения очень высока, покупатели будут более заинтересованы в анализе сигналов и с большей вероятностью выберут сигналы с высокой прогностической ценностью и низкой достоверностью.
Таким образом, риск неправильной интерпретации сигналов с высокой прогностической ценностью/низкой достоверностью воспринимается как меньший, а потенциальная выгода в плане того, что выбор не будет ошибочным, увеличивается, в результате чего покупатели больше зависят от высокой прогностической ценности/низкой достоверности ценностные ориентиры.
МЕТОД
Исследование проводилось в два этапа. Первый этап включал в себя три опроса, измеряющих предполагаемую прогностическую и доверительную ценность сигналов, которые можно было бы использовать при формировании суждения об относительном качестве различных видов пейджеров (или «пейджеров», как их иногда называют). Вопросы были основаны на тех, которые использовал Олсон (Olson and Jacoby, 1972). Чтобы измерить прогностическую ценность сигнала, испытуемые реагировали по шкале от одного до пяти. с одним пометкой «совсем не точно» и пятью с пометкой «чрезвычайно точно», насколько, по их мнению, «следующие типы информации точно указывают на общее качество ранда бумаги?» Затем испытуемые указали свою воспринимаемую ценность уверенности в том же наборе сигналов. Субъекты ответили по шкале от одного до пяти, где один отмечен как «совсем не уверен», а пять отмечены как «чрезвычайно уверен», насколько они уверены, что знают, «что означает каждый из следующих терминов?»
Субъекты ответили по шкале от одного до пяти, где один отмечен как «совсем не уверен», а пять отмечены как «чрезвычайно уверен», насколько они уверены, что знают, «что означает каждый из следующих терминов?»
Первые два опроса определили список из восьми сигналов, которые можно было использовать на втором этапе, четыре из которых были восприняты как имеющие высокую прогностическую ценность, но низкую доверительную ценность, и четыре, которые были восприняты как низкие прогностическая ценность и высокая достоверность. В финальном опросе приняли участие 50 испытуемых из выборки (учащиеся, зачисленные на вводный курс по организационному поведению), чтобы подтвердить воспринимаемую ценность достоверности и прогностическую ценность этих сигналов, см. рис. 1).
Во второй фазе использовался факторный эксперимент 2 X 2 с двумя уровнями нехватки времени и двумя уровнями риска. Пятьдесят два испытуемых участвовали в каждом из четырех возможных условий.
В эксперименте одновременно участвовало до восьми испытуемых. Они сидели за отдельными столиками, у каждого по одной-две брошюры из картонных страниц с восемью вкладками на каждой странице. На каждой странице было восемь подсказок о бренде, причем бренды были помечены только цифрами 1–5. Каждая вкладка была помечена как подсказка, например «стабильность частоты», и испытуемым сказали, что они могут узнать информацию об этом конкретном бренде, потянув за вкладку и прочитав, что написано под ней.
Они сидели за отдельными столиками, у каждого по одной-две брошюры из картонных страниц с восемью вкладками на каждой странице. На каждой странице было восемь подсказок о бренде, причем бренды были помечены только цифрами 1–5. Каждая вкладка была помечена как подсказка, например «стабильность частоты», и испытуемым сказали, что они могут узнать информацию об этом конкретном бренде, потянув за вкладку и прочитав, что написано под ней.
Само задание заключалось в том, что испытуемые слушали короткую лекцию об истории пейджеров и о том, как они работают. Затем они индивидуально отбирали сигналы, выбирая только четыре из восьми сигналов, доступных для каждого бренда, а затем записывали на отдельной странице порядок, в котором они выбирали сигналы по мере перехода от бренда один к бренду пять. Выбрав свои реплики, они могли вернуться и проанализировать реплики, если хотели, а затем выбрать бренд, который, по их мнению, имел самое высокое качество. Поскольку им не сообщали цены на альтернативные бренды, они не могли оценить относительную стоимость, и поэтому их не просили выбрать пейджер, который они хотели бы купить. Чтобы дополнительно контролировать любые ценовые эффекты, например, если испытуемые предполагают, что пейджеры mate на Тайване дешевле, чем пейджеры mate в L’.S.A., им заранее сказали, что все бренды оцениваются приблизительно в 9 раз.0003
Чтобы дополнительно контролировать любые ценовые эффекты, например, если испытуемые предполагают, что пейджеры mate на Тайване дешевле, чем пейджеры mate в L’.S.A., им заранее сказали, что все бренды оцениваются приблизительно в 9 раз.0003
РИСУНОК 1
ОТНОСИТЕЛЬНОЕ ПОЛОЖЕНИЕ ПЯТНАДЦАТИ ПРИЗНАКОВ НА ГРАФИКЕ В СООТВЕТСТВИИ С ИХ СРЕДНИМИ ПРОГНОЗИРУЕМЫМИ ЗНАЧЕНИЯМИ (ГОРИЗОНТАЛЬНАЯ ОСИ) И СРЕДНИМИ ДОВЕРИТЕЛЬНЫМИ ЗНАЧЕНИЯМИ (ВЕРТИКАЛЬНАЯ ОСИ).
Пейджеры были выбраны потому, что это продукт, с которым мало кто знаком. Это важно по двум причинам. Во-первых, знакомство с классом продукта может быть важным фактором, определяющим выбор и использование сигнала. Используя продукт, с которым никто не знаком (или устраняя тех испытуемых, которые с ним знакомы), знакомство контролируется в эксперименте.
Во-вторых, чтобы манипулировать предполагаемым риском, необходимо было привести две очень разные истории о производительности и средней цене различных брендов, доступных для потребителей. Если испытуемые не были знакомы с продуктом, было мало шансов, что они обнаружат обман. Для дальнейшего контроля узнаваемости торговые марки оцениваемых пейджеров не были включены в набор признаков. Некоторые испытуемые, возможно, знакомы с этими торговыми марками, возможно, даже имея опыт работы с другими продуктами того же производителя.
Если испытуемые не были знакомы с продуктом, было мало шансов, что они обнаружат обман. Для дальнейшего контроля узнаваемости торговые марки оцениваемых пейджеров не были включены в набор признаков. Некоторые испытуемые, возможно, знакомы с этими торговыми марками, возможно, даже имея опыт работы с другими продуктами того же производителя.
Еще одной причиной использования пейджеров было то, что продукт носит технический характер. Таким образом, разумно предположить, что испытуемые хотели бы использовать технические подсказки для оценки брендов. Предполагалось, что эти сигналы будут рассматриваться испытуемыми как имеющие высокую прогностическую ценность и низкую достоверность.
Главной зависимой переменной в этом исследовании было количество сигналов с высокой достоверностью и низкой прогностической ценностью, полученных субъектом. Испытуемый пикетирует четыре признака для каждого из пяти брендов, что дает переменной диапазон от 0 до 20. Основной зависимой переменной могло бы в равной степени быть количество сигналов с низким уровнем достоверности и высокой прогностической ценностью, поскольку сумма двух типов используемых сигналов будет всегда равно двадцати.
Основная зависимая переменная измеряла выбор подсказки, поскольку выбор подсказки был в центре внимания исследования. Это произошло потому, что анализ выбора сигналов был логическим первым шагом к анализу роли сигналов в процессе принятия решений. Кроме того, с точки зрения маркетолога или потребителя, заставить покупателя обращать внимание на определенные сигналы является необходимым первым шагом к тому, чтобы повлиять на выбор потребителя.
Однако вместо того, чтобы позволить испытуемым просто выбирать подсказки, было решено, что они должны продолжить процесс выбора, оценивая бренды и выбирая бренд, который, по их мнению, был лучшим. Затем анализ мог бы изучить возможное влияние выбора сигнала и условий эксперимента на использование сигналов после того, как они были выбраны. Поскольку этот анализ был добавлен в исследовательских целях, перед экспериментом не было выдвинуто ни одной гипотезы. Однако было сочтено, что нехватка времени и предполагаемый риск могут дополнительно влиять на использование реплик, так что эффект выбора реплики увеличивается или уменьшается.
Сигналы, которые люди фактически используют для оценки продукта, в прошлых исследованиях были идентифицированы путем регрессии значений сигналов с оценками объектов субъектом, чтобы определить, какие сигналы предсказывают рейтинги (например, Slovic and MacPhillany, 1974). Это было невозможно в настоящей задаче, поскольку не было достаточного количества наблюдений для проведения регрессионного анализа, особенно потому, что многие значения сигналов были категориальными и должны были быть преобразованы в фиктивные переменные. Вместо этого сразу после выбора марки пейджера, которую они считали наиболее качественной, испытуемых просили проанализировать свой мыслительный процесс и перечислить сигналы, которые они фактически использовали при принятии решения. Второй набор зависимых переменных был получен из этого вопроса. Одним из них было количество сигналов с высокой степенью достоверности и низкой прогностической ценностью, которые, по их словам, они фактически использовали при выборе бренда, а другим — количество признаков с высокой прогностической ценностью и низкой достоверностью, которые они использовали. Эти переменные теоретически могут варьироваться от нуля до четырех для обоих типов сигналов для каждого субъекта. Однако, в отличие от основной зависимой переменной, приведенной выше, их сумма не обязательно должна быть постоянной, поэтому в ходе анализа рассматривались обе переменные.
Эти переменные теоретически могут варьироваться от нуля до четырех для обоих типов сигналов для каждого субъекта. Однако, в отличие от основной зависимой переменной, приведенной выше, их сумма не обязательно должна быть постоянной, поэтому в ходе анализа рассматривались обе переменные.
Третьей зависимой переменной была торговая марка, признанная самой качественной. Значения сигналов были такими, что наилучший выбор с использованием сигналов с высокой прогностической ценностью отличался от выбора с использованием сигналов с высокой достоверностью. Если бы гипотезы были верны, распределение брендов, оцененных как наиболее качественные, изменилось бы в предсказуемом направлении. Например, пейджер номер три имел хорошие отзывы пользователей, был произведен в Англии (в отличие от Тайваня и Японии), продавался компанией Sears (в отличие от Zayre и Ayr-Way) и имел наибольшую долю рынка. «С другой стороны, у него был самый низкий частотный диапазон, худшая чувствительность к поисковым вызовам, вторая худшая селективность по соседнему каналу и он был наравне с четырьмя другими брендами по стабильности частоты. Таким образом, субъекты, использующие сигналы с высокой прогностической ценностью, не должны были пикетировать этот бренд, в то время как субъекты, использующие сигналы с высокой достоверностью, должны были чаще его выбирать.
Таким образом, субъекты, использующие сигналы с высокой прогностической ценностью, не должны были пикетировать этот бренд, в то время как субъекты, использующие сигналы с высокой достоверностью, должны были чаще его выбирать.
Если бы гипотезы были верны, третий пейджер следовало бы чаще выбирать в условиях высокой нехватки времени и низкого риска. Поскольку это была косвенная мера влияния манипуляций на выбор реплики, она была зарезервирована как вторичная зависимая переменная. Однако было сочтено, что выводы можно было бы придать гораздо большей важности, если бы было показано, что манипуляция не только изменила поведение при выборе сигнала, но и вызвала изменение суждения, вынесенного субъектом.
Нехватка времени была манипулирована за счет того, что группа с высокой степенью нехватки времени оценивала пейджеры, а затем камеры (используя аналогичный буклет) в то время, когда группа с низкой нехваткой времени оценивала пейджеры. Оценка камер после пейджеров гарантировала, что группа спешки выполнила свою задачу в отведенное время. Предварительные тесты использовались для определения оптимального времени для двух уровней лечения на основе разброса баллов в проверках манипулирования и обсуждения с участниками их реакции на Давление времени.
Предварительные тесты использовались для определения оптимального времени для двух уровней лечения на основе разброса баллов в проверках манипулирования и обсуждения с участниками их реакции на Давление времени.
Риском манипулировали, заставляя группы с высоким уровнем риска прочитать цитату из Consumer’s Reports и другую цитату из правительственного исследования (оба вымышленные), в которых говорится, что все пейджеры, как правило, хорошо работают в тестах и что шансы купить тот, который не работает ну минимальны. Группе высокого риска было сказано, что несколько брендов не работают хорошо, и шансы купить неудовлетворительный бренд весьма значительны. Достоверность и влияние этих отчетов были проверены в ходе предварительных испытаний.
Риском также манипулировали, сообщая испытуемым, что пейджеры стоят около 20 долларов в ситуации низкого риска и около 450 долларов в ситуации высокого риска.
РЕЗУЛЬТАТЫ
Проверки манипулирования показали сильные различия в восприятии нехватки времени (p c. 0001) и предполагаемого риска (p <.0001). Основные эффекты нехватки времени для выбора подсказки были значительными, F = 5,51 (p = 0,02), а взаимодействие нехватки времени и риска было погранично значимым при F = 3,73, p = 0,055 (см. Рисунок 2). Основные эффекты риска не были значительными. Тест Тьюки HSD между средними значениями показал значительную разницу между средним значением для состояния с высоким давлением времени и низким риском (X = 10,48) и состоянием с низким давлением времени и низким риском (X = 7,9).) при р<0,05. Общая объясненная дисперсия составила W2=4,18.
0001) и предполагаемого риска (p <.0001). Основные эффекты нехватки времени для выбора подсказки были значительными, F = 5,51 (p = 0,02), а взаимодействие нехватки времени и риска было погранично значимым при F = 3,73, p = 0,055 (см. Рисунок 2). Основные эффекты риска не были значительными. Тест Тьюки HSD между средними значениями показал значительную разницу между средним значением для состояния с высоким давлением времени и низким риском (X = 10,48) и состоянием с низким давлением времени и низким риском (X = 7,9).) при р<0,05. Общая объясненная дисперсия составила W2=4,18.
РИСУНОК 2
СРЕДНЕЕ КОЛИЧЕСТВО РЕКОМЕНДАЦИЙ С ВЫСОКИМ CV
РИСУНОК 3 субъекты сказали, что они использовали в решении. Основные эффекты нехватки времени были очень значимыми (p<0,001), при этом основной эффект риска и эффект взаимодействия существенно не различались (см. рис. 3). Объясненная дисперсия была выше для этой зависимой переменной при w2=0,086.
Третьей зависимой переменной был пейджер, признанный брендом самого высокого качества. Распределение брендов значительно отличалось между условиями высокого и низкого давления времени X2 = 10,35, p = 0,025 (см. Таблицу 1). Ожидается, что субъекты, зависящие от сигналов с высокой достоверностью/низкой прогностической ценностью, будут чаще выбирать третью марку. Как и предсказывалось, процент выбора этой марки вырос с 42,9% до 62,2% при переходе от условий низкой нехватки времени к условиям высокой нехватки времени (p-0,005). Ожидается, что субъекты, зависящие от сигналов с низкой достоверностью и высокой прогностической ценностью, будут чаще выбирать пятую марку. Как и предполагалось, процент испытуемых, выбравших этот бренд, вырос с 14,4% до 25,9%.% от состояния высокого цейтнота до состояния низкого цейтнота (p-0,025).
ОБСУЖДЕНИЕ
Нехватка времени, по-видимому, влияет на выбор человеком сигналов и его зависимость от выбранных сигналов. Было продемонстрировано, что кумулятивный эффект вызывает изменение суждений человека. Субъекты в условиях сильного цейтнота, которые указывали на то, что они испытали значительный цейтнот, были теми, кто, как правило, не использовал сигналы с высокой достоверностью (v-9)..13). Те, кто указал, что испытывал небольшой дефицит времени, больше зависели от сигналов с высокой достоверностью (х-12,29). Это говорит о том, что использование подсказок с высокой достоверностью воспринимается как стратегия экономии времени, вызывающая изменение выбора подсказки и поведения при использовании подсказки.
Было продемонстрировано, что кумулятивный эффект вызывает изменение суждений человека. Субъекты в условиях сильного цейтнота, которые указывали на то, что они испытали значительный цейтнот, были теми, кто, как правило, не использовал сигналы с высокой достоверностью (v-9)..13). Те, кто указал, что испытывал небольшой дефицит времени, больше зависели от сигналов с высокой достоверностью (х-12,29). Это говорит о том, что использование подсказок с высокой достоверностью воспринимается как стратегия экономии времени, вызывающая изменение выбора подсказки и поведения при использовании подсказки.
Этот эффект мог бы быть еще более выраженным, если бы сигналы характеризовались количеством когнитивных усилий, необходимых для принятия решения о соответствующей категории объекта по интересующему атрибуту, и эта мера использовалась бы в качестве зависимой переменной, а не полученной доверительное значение сигнала. 9Таблица 1 В этом эксперименте у испытуемых было два типа сигналов на выбор. Они могли выбрать сигналы, которые, по их мнению, были менее предсказуемы в отношении качества, но сигналы, которые, как они были уверены, они поняли. Понимание сигнала, по-видимому, позволит им лучше оценить объект. приводит к лучшему выбору.
Они могли выбрать сигналы, которые, по их мнению, были менее предсказуемы в отношении качества, но сигналы, которые, как они были уверены, они поняли. Понимание сигнала, по-видимому, позволит им лучше оценить объект. приводит к лучшему выбору.
Второй стратегией будет выбор сигналов, которые, по их мнению, имеют высокую прогностическую ценность, но которые, как они не уверены, понимают. Это заставило бы их каким-то образом узнать, что было или не было хорошей ценностью для конкретного объекта. Они никогда не могли быть уверены, правильно ли они классифицируют объект, но чувствовали, что их окончательный выбор был лучше, поскольку он был основан на признаках, которые, по их мнению, были лучшими предикторами качества.
В ходе эксперимента оказалось, что испытуемые предпочитали больше полагаться на последнюю стратегию, даже в ситуации с низким риском. «Только введение значительного дефицита времени вынудило их использовать подсказки с высокой достоверностью в качестве стратегии экономии времени.
Величина дисперсии, объясняемая нехваткой времени и манипулированием рисками, невелика. Тем не менее, похоже, что объясненная дисперсия недооценивает степень влияния нехватки времени на процесс, поскольку доля людей, оценивающих бренды три и пять как наилучшие, резко менялась в зависимости от условий. Если бы это можно было перевести непосредственно в доли рынка (при условии, что люди покупают марку, которую они считают самой качественной), влияние на фирмы, продающие марки 3 и 5, было бы огромным. Эти результаты свидетельствуют о значительном влиянии поведения при выборе подсказки на суждения потребителей. До тех пор, пока не будут поняты детерминанты выбора и использования сигналов, большая часть вариаций в процессе принятия решений потребителем не будет объяснена. Было обнаружено, что нехватка времени и, в меньшей степени, предполагаемый риск являются определяющими факторами выбора сигнала.
ССЫЛКИ
Эндрюс, И. Р. и Валенци, Э. Р. (1971), Сочетание цен, торговых марок и признаков магазина для формирования впечатления о качестве продукта», Труды, 79-я ежегодная конвенция, АПА, 649-50.
Р. (1971), Сочетание цен, торговых марок и признаков магазина для формирования впечатления о качестве продукта», Труды, 79-я ежегодная конвенция, АПА, 649-50.
Арчер, Э. Джеймс (1962), Идентификация понятий как функция очевидности релевантной и нерелевантной информации, Журнал экспериментальной психологии, 63, 6, 616-620.
Беттман, Дж. Р. и Джейкоби, Джейкоб (1978), «Модели обработки информации о потребителях, достижения в исследованиях потребителей», 3, 15–20.
Брим, О. Г. младший (1954), «Принятие нового поведения в воспитании детей, человеческие отношения», 7, 473-491.
Брунер, Дж. С., Гуднау, Джекелин Дж. и Остин, Г. А. (1956), Исследование мышления, Нью-Йорк: Wiley,
Брансуик, Э. (1949), Систематический и репрезентативный план психологических экспериментов, Беркли: University of California Press,
Кокс, Д. Ф. (1967), Риск-принятие и информация- Управление потребительским поведением, Бостон, издательство Гарвардского университета, стр. 9.0003
9.0003
Эйнхорн, Х. Дж., Клейнмерц, Д. Н. и Клейнмерц, Б. (1979), Линейная регрессия и модели суждения с отслеживанием процессов, «Психологический обзор», 86, 465-85.
Френч, Дж. Р. П., младший и Снайдер, Р. (1959), Лидерство и межличностная власть, в Д. Картрайт (ред.), Исследования социальной власти, Анн-Арбор: Мичиганский университет Press, 118-149.
Ховард, Джон А. и Шет, Джагдиш Н. (1969), The Theory of Buyer Behavior, New York: Wiley,
Jacoby, Jacob A., Chestnut, Robert A., Weigl, Karl C. and Fisher, William (1975), «Сбор информации перед покупкой: описание методологии процесса, исследовательская парадигма и пилотное исследование», Беверли Б. Андерсон (ред.), «Достижения в исследованиях потребителей», 3, 306-16
Джейкоби, Джейкоб, Сибилло, Джордж Дж. и Бузато-Шах, Жаклин (1977), «Поведение при сборе информации в ситуациях выбора бренда», Журнал потребительских исследований, 3 (март) 209–16.
Келли, Х. Х. и Арровуд, А. Дж. (1960) , «Коалиции в триаде: критика и эксперимент», Sociometry, 23, 231–44.
Кон, Кэрол А. и Джейкоби, Джейкоб (1973), «Модели получения информации при покупке новых продуктов», в Скотт Уорд и Питер Райт, (ред.), Достижения в исследованиях потребителей, 1, 427-30.
Олсон, Джерри С. и Джейкоби, Джейкоб (1972), «Использование сигналов в процессах восприятия качества», М. Венкатесан, (ред.), Ассоциация потребительских исследований, Труды, 167–79.
Пинкус, Стивен и Л.К. Уотерс (1975), Рейтинги качества продукции как функция наличия внутренних признаков продукта и информации о цене, Журнал прикладной психологии, 60, 280-82.
Роллингс, Гарри; Бетел, Барбара и Деффенбакер, Кеннет (1971), Соответствующие эффекты размещения реплик в задачах идентификации концепций, Журнал экспериментальной психологии, № 7, 1, 9-12.
Сарбин, Т. Р.; Тафт, Р. и Бейли, Д. Э. (1960), Клинический вывод и когнитивная теория, Нью-Йорк: Холт, Райнхарт и Уинстон.
Э. (1960), Клинический вывод и когнитивная теория, Нью-Йорк: Холт, Райнхарт и Уинстон.
Шеллинк, Дуглас А. (1980), Детерминанты поведения при выборе сигнала, неопубликованная диссертация, Университет Иллинойса.
Словик, Пол и Макфилани, Дуглас (1974), «Соизмеримость измерений и использование подсказок в сравнительном суждении», Organizational Behavior and Human Performance, II, 172-194.
Тверски, А. и Канеман, Д. (1980), «Формирование решений и рациональность». по выбору, факультет психологии, Стэнфордский университет.
Винаке, У. Э. и Аркофф, А. (1957), Экспериментальное исследование коалиций в триаде, «American Sociological Review», 22, 406-14.
————— —————————
Характеристики волн около берега в качестве сигналов для ориентации на плавание в пружине флагэк -черепахи
ScienceDirectКорпоративный знак.
https://doi.org/10.1016/j. jembe.2020.151475Получить права и контент
jembe.2020.151475Получить права и контент
Считается, что волны служат важным ориентиром для вылупившихся морских черепах при перемещении через прибрежную зону и облегчении распространения в океанских водах. . Как плоскоспинная черепаха ( Natator depressus ) — единственный вид морских черепах, у которого отсутствует океаническая ювенильная стадия и который остается на континентальном шельфе на протяжении всего жизненного цикла, возможно, что вылупившиеся детеныши этого вида не используют волновые сигналы для раннего расселения. Здесь мы использовали волновой лоток, чтобы изучить реакцию вылупившихся детенышей плоскоспинных черепах на волны как сигнал для рассредоточения у берега. Мы подвергали вылупившихся птенцов воздействию двух типов волн (морских волн и зыби), различающихся по высоте и периоду, которые обычно наблюдаются в местах гнездования, и следили за направлением плавания (ориентацией). Птенцы ориентированы на морские волны с более коротким периодом (3 с) как с небольшой (6 см от пика до впадины), так и с большой (12 см от пика до впадины) высотой волн. Ориентация вылупившихся детенышей на волны зыби с более длительным периодом (8 с) происходила только при больших (16 см от пика до впадины), а не при малых (7 см от пика до впадины) высотах волн. Ускорение, создаваемое волнами, было самым сильным предиктором предпочтения направления у вылупившихся осанок. Наблюдалась положительная связь между максимальным ускорением и предпочтением направления, при этом вылупившиеся осанки были более концентрированными (значение r 0,9).8) в направлении набегающей волны с волнами, дающими наибольшее ускорение (морские волны с наибольшей высотой волн). Эти волны были похожи на морские волны, генерируемые прибрежными ветрами, которые были наиболее распространенным типом волн, которые мы измеряли на пляже, где гнездятся плоскоспинные черепахи. Наше исследование подтвердило, что, несмотря на отсутствие океанической стадии развития, плоскоспинные детеныши, как и другие виды морских черепах, могут обнаруживать сигналы волн и реагировать на них, и что ускорение, создаваемое волнами, может быть ключевой характеристикой волн, определяющей ориентацию.
Ориентация вылупившихся детенышей на волны зыби с более длительным периодом (8 с) происходила только при больших (16 см от пика до впадины), а не при малых (7 см от пика до впадины) высотах волн. Ускорение, создаваемое волнами, было самым сильным предиктором предпочтения направления у вылупившихся осанок. Наблюдалась положительная связь между максимальным ускорением и предпочтением направления, при этом вылупившиеся осанки были более концентрированными (значение r 0,9).8) в направлении набегающей волны с волнами, дающими наибольшее ускорение (морские волны с наибольшей высотой волн). Эти волны были похожи на морские волны, генерируемые прибрежными ветрами, которые были наиболее распространенным типом волн, которые мы измеряли на пляже, где гнездятся плоскоспинные черепахи. Наше исследование подтвердило, что, несмотря на отсутствие океанической стадии развития, плоскоспинные детеныши, как и другие виды морских черепах, могут обнаруживать сигналы волн и реагировать на них, и что ускорение, создаваемое волнами, может быть ключевой характеристикой волн, определяющей ориентацию.
Молодые морские животные используют различные физические сигналы, такие как течения, ветер и волны, чтобы облегчить расселение из естественной среды обитания (Able, 1991; Chapman et al., 2011; Huijbers et al., 2012; Jeffs et al., 2005). . Определение относительной важности отдельных сигналов для паттернов движения может быть сложной задачей и обычно требует экспериментального подхода. Эта информация имеет решающее значение для управления, поскольку она может помочь в выявлении и смягчении антропогенных угроз, которые могут нарушить перемещения этих уязвимых этапов жизненного цикла (Kokko and Lopez-Sepulcre, 2006). Эти угрозы могут включать в себя прибрежные сооружения, такие как дамбы или волнорезы, которые могут изменить характер волн или движение океанских течений, если они расположены рядом с пляжами.
Детеныши морских черепах вылупляются из гнезд на пляжах и направляются к океану, используя визуальные ориентиры, двигаясь к более яркому нижнему горизонту и удаляясь от приподнятых темных силуэтов (Limpus and Kamrowski, 2013; Mrosovsky and Shettleworth, 1968; Salmon et al. ., 1992). Добравшись до моря, многие виды непрерывно плавают в течение как минимум 24 часов, что приводит к быстрому перемещению от мелководных местообитаний, где много хищников, к относительно безопасным более глубоким водам (Дрофа, 1972; Wyneken and Salmon, 19).92). Считается, что они используют направление волны в качестве ориентира для перемещения в море (Ломанн и Ломанн, 1992; Ломанн и др., 1990; Ломанн и др., 1995; Окуяма и др., 2009; Винекен и др., 1990). обнаружение орбитального движения волн для ориентации в направлении волны (Lohmann et al., 1995). Это поведение не является визуальным, поскольку лабораторные исследования в аквариумах с волнами показали, что вылупившиеся детеныши могут обнаруживать волны в отсутствие света (Lohmann et al., 1990; Wyneken et al., 1990), хотя зрение может улучшить их способность или мотивацию к реагированию. (Гофф и др., 1998). Лабораторные эксперименты показали, что вылупившиеся детеныши используют для навигации магнитное поле Земли с помощью волновой ориентации, чтобы ориентироваться в море (Goff et al.
., 1992). Добравшись до моря, многие виды непрерывно плавают в течение как минимум 24 часов, что приводит к быстрому перемещению от мелководных местообитаний, где много хищников, к относительно безопасным более глубоким водам (Дрофа, 1972; Wyneken and Salmon, 19).92). Считается, что они используют направление волны в качестве ориентира для перемещения в море (Ломанн и Ломанн, 1992; Ломанн и др., 1990; Ломанн и др., 1995; Окуяма и др., 2009; Винекен и др., 1990). обнаружение орбитального движения волн для ориентации в направлении волны (Lohmann et al., 1995). Это поведение не является визуальным, поскольку лабораторные исследования в аквариумах с волнами показали, что вылупившиеся детеныши могут обнаруживать волны в отсутствие света (Lohmann et al., 1990; Wyneken et al., 1990), хотя зрение может улучшить их способность или мотивацию к реагированию. (Гофф и др., 1998). Лабораторные эксперименты показали, что вылупившиеся детеныши используют для навигации магнитное поле Земли с помощью волновой ориентации, чтобы ориентироваться в море (Goff et al. , 1998; Lohmann and Lohmann, 1996; Lohmann et al., 1997). Такие исследования в основном были сосредоточены на волнах с коротким периодом <2 с (Ломанн и др., 1995; Винекен и др., 1990), но не определяли точных характеристик, таких как период, высота, ускорение и крутизна, на которых можно ориентироваться. быть зависимым. Симулятор волн, который использует механическую руку для имитации в воздухе орбитального движения волн, которое детеныш испытал бы в воде (Lohmann et al., 19).95) продемонстрировал, что вылупившиеся болванчики ( Caretta caretta ) могут обнаруживать волны разного периода и высоты и что они, по-видимому, реагируют на тип волны, с которой столкнулись на родине (Manning et al., 1997).
, 1998; Lohmann and Lohmann, 1996; Lohmann et al., 1997). Такие исследования в основном были сосредоточены на волнах с коротким периодом <2 с (Ломанн и др., 1995; Винекен и др., 1990), но не определяли точных характеристик, таких как период, высота, ускорение и крутизна, на которых можно ориентироваться. быть зависимым. Симулятор волн, который использует механическую руку для имитации в воздухе орбитального движения волн, которое детеныш испытал бы в воде (Lohmann et al., 19).95) продемонстрировал, что вылупившиеся болванчики ( Caretta caretta ) могут обнаруживать волны разного периода и высоты и что они, по-видимому, реагируют на тип волны, с которой столкнулись на родине (Manning et al., 1997).
Плоская черепаха — единственный вид морских черепах, у которого нет пелагической фазы; но остается на континентальном шельфе в неритовой зоне (Bolten, 2003; Walker, 1991; Walker, Parmenter, 1990). Учитывая другой образ жизни и тот факт, что они иногда гнездятся на более защищенных пляжах, чем зеленые черепахи (9). 0111 Chelonia mydas ), где поверхностные волны в основном генерируются местными ветрами, а не океанической зыбью (Pendoley, 2005), неясно, будут ли они вести себя так же, как другие виды черепах, и будут ли они ориентироваться на фронт волн. Здесь мы использовали волновой лоток, чтобы проверить, могут ли детеныши плоскоспинных черепах ориентироваться в волнах, и если да, то определить характеристики волн, которые вызвали самую сильную реакцию. Мы использовали характеристики волн (высота, период), измеренные на пляже, где гнездятся плоскоспинные черепахи, чтобы настроить ряд реалистичных экспериментальных волновых процедур в волновом лотке (подробности см. В дополнительных данных). Мы предположили, что в отсутствие каких-либо других сигналов вылупившиеся детеныши будут ориентироваться на фронт волны, когда волны присутствуют. Кроме того, поскольку этот вид гнездится на незащищенных от ветра пляжах, где, вероятно, преобладают морские волны (генерируемые местными ветрами) (период волн <6 с) (Santana Garcon et al.
0111 Chelonia mydas ), где поверхностные волны в основном генерируются местными ветрами, а не океанической зыбью (Pendoley, 2005), неясно, будут ли они вести себя так же, как другие виды черепах, и будут ли они ориентироваться на фронт волн. Здесь мы использовали волновой лоток, чтобы проверить, могут ли детеныши плоскоспинных черепах ориентироваться в волнах, и если да, то определить характеристики волн, которые вызвали самую сильную реакцию. Мы использовали характеристики волн (высота, период), измеренные на пляже, где гнездятся плоскоспинные черепахи, чтобы настроить ряд реалистичных экспериментальных волновых процедур в волновом лотке (подробности см. В дополнительных данных). Мы предположили, что в отсутствие каких-либо других сигналов вылупившиеся детеныши будут ориентироваться на фронт волны, когда волны присутствуют. Кроме того, поскольку этот вид гнездится на незащищенных от ветра пляжах, где, вероятно, преобладают морские волны (генерируемые местными ветрами) (период волн <6 с) (Santana Garcon et al.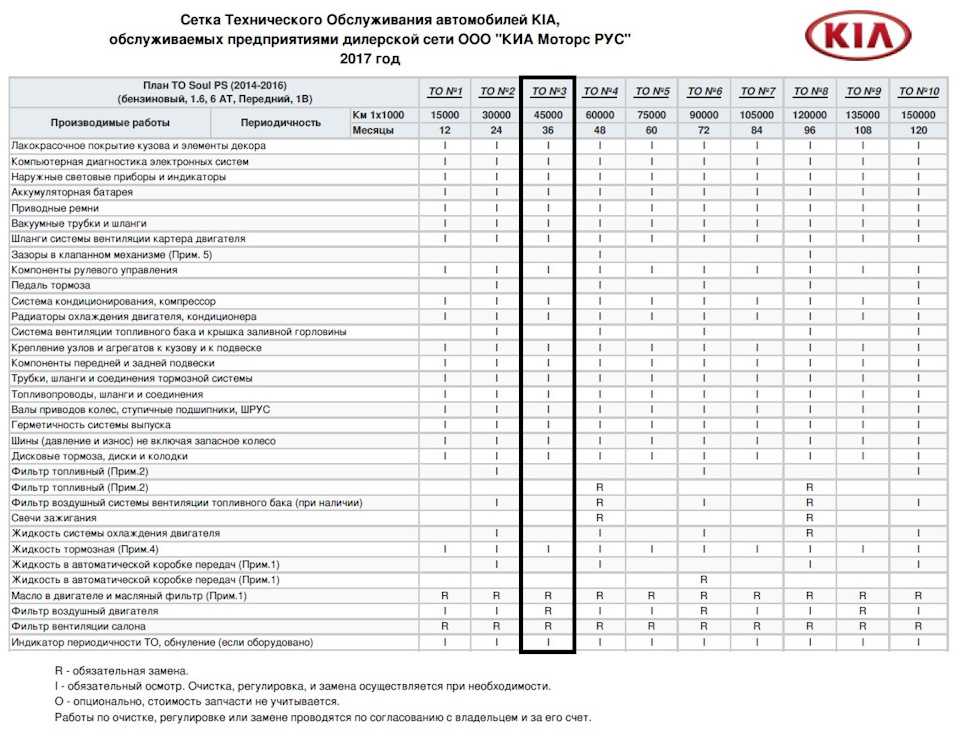 , 2010), реакция вылупившихся детенышей на короткопериодные волны, обычно создаваемые ожидается, что ветер будет более выраженным, чем реакция на волны зыби с более длительным периодом (период волны ≥8 с).
, 2010), реакция вылупившихся детенышей на короткопериодные волны, обычно создаваемые ожидается, что ветер будет более выраженным, чем реакция на волны зыби с более длительным периодом (период волны ≥8 с).
Фрагменты срезов
Птенцы ( n = 55) были получены с пляжа Беллс-Бич, материкового места гнездования плоскоспинных черепах в районе Пилбара в Западной Австралии, длиной примерно 550 м и расположенного между 20°36′49″ ю.ш. 117°09′03″ в.д. и 20°36′32″ ю.ш., 117°09′28″ в.д. (рис. S.1). Ночью на пляже наблюдали за следами только что вылупившихся детенышей. Когда были обнаружены следы, их отслеживали обратно в гнездо, а птенцов собирали либо по мере их появления, либо с вершины гнезда. Они были помещены в
Сочетание черной пластиковой рамы и экспериментов, проводимых ночью, привело к тому, что птенцы оказались в полной темноте во время экспериментов в лотке. Температура воды в лотке 26 января 30°С, 29 января 28°С. Средняя глубина воды (± SD), измеренная датчиком давления, составила 0,62 ± 0,03 м. Максимальная H с (рис. S.3), T и длина волны (L), крутизна волны (s), максимальная горизонтальная скорость поверхностной воды и максимальное ускорение в каждой обработке
Максимальная H с (рис. S.3), T и длина волны (L), крутизна волны (s), максимальная горизонтальная скорость поверхностной воды и максимальное ускорение в каждой обработке
Наш эксперимент с большим волновым лотком, способным создавать волны разной высоты и периодов, показал, что вылупившиеся детеныши плоскоспинных черепах ориентируются на различные волны, характерные для тех, которые возникают в местах их гнездования, и по проверенным параметрам предпочтение отдается направлению волн. преимущественно связано с максимальным ускорением волны, функцией крутизны волны, высоты волны и периода волны, с максимальной ориентацией ( r = 0,98), когда присутствуют большие морские волны.
П.В. был поддержан стипендией Австралийской правительственной программы подготовки научных кадров (RTP) и дополнительной стипендией Университета Западной Австралии. Финансирование проекта было предоставлено в основном Департаментом биоразнообразия, охраны природы и достопримечательностей в рамках Программы сохранения плоскоспинных черепах Северо-Западного шельфа при участии всех организаций-авторов.
Все авторы задумали и разработали эксперименты; П.В., К.П., К.П., М.Т., С.В. провел полевые работы и провел лотковые эксперименты; П.В. выполнил статистический анализ по совету М.Т.; П.В. подготовил рукопись, и все авторы внесли правки.
Авторы заявляют об отсутствии конфликта интересов.
Особая благодарность членам программы West Pilbara Turtle Program, особенно Кариссе Айвз, а также Джейсону Россенделлу из Rio Tinto за их помощь на Беллс-Бич. Также выражаем благодарность Брэду Роузу, Дайанн Кинг, Джеймсу Хере, Арнольду ван Ройену, Мариам Абдолахпур, Блер Бентли, Николе Митчелл, Селине Уорбертон, Минь Во, Лусиане Феррейре, Данниэль Пилчер, Грэму Уилсону, Кристе Ханнан, Джессике Райли и Каприс Хайд (DBCA). ) за помощь. Спасибо Деннису Стэнли за помощь с ИК-камерой
Ссылки (58)
- J. Wyneken et al.
Ориентация вылупившихся морских черепах
Caretta caretta L. в аквариуме с искусственными волнамиJ. Exp.
 Мар биол. Экол.
Мар биол. Экол.(1990)
- П. Уилсон и др.
Массовое хищничество детенышей морских черепах вблизи прибрежной пристани
Biol. Консерв.
(2019)
- М. Лосось и др.
Ориентация и плавание вылупившихся черепах логгерхедов
Caretta caretta L. во время морской миграцииJ. Exp. Мар биол. Экол.
(1987)
- Э.С. Полочанска и др.
Уязвимость морских черепах к изменению климата
Доп. Мар биол.
(2009)
- К. Б. Паттиаратчи и др.
Воздействие морского бриза на прибрежные и прибрежные процессы на юго-западе Австралии
Прод. Полка Рез.
(1997)
- Дж. Окуяма и др.
Онтогенез дисперсионной миграции зеленой черепахи (
Chelonia mydas ) вылупившихся детенышейJ. Exp. Мар биол. Экол.
(2009)
- К. Ф. Nordstrom
Жизнь с берегоукрепительными сооружениями: обзор
Устье. Побережье. Шельф науч.
(2014)
- A. Harewood et al.
Воздействие прибрежной застройки на выживаемость детенышей бисса и успех плавания во время первоначальной миграции в открытом море
Биол. Консерв.
(2008)
- М. Гофф и др.
Детеныши морских черепах используют поверхностные волны для определения направления по магнитному компасу
Аним. Поведение
(1998)
- Дж.В. Чепмен и др.
Стратегии ориентации животных для движения в потоках
Curr. биол.
(2011)
 Мар биол. Экол.
Мар биол. Экол. Ф. Nordstrom
Ф. NordstromНаблюдения и моделирование морского бриза: обзор
австр. Метерол. Магаз.
(1992)
Общие темы и вариации в системах ориентации животных
Am. Зоол.
(1991)
 Agostinelli
AgostinelliCircular Statistics, from «Topics in Circular Statistics» (2001)
R package Version 0.2-6
(2018)
Динамика побережья с системой волноломов, Двенадцатая конференция по береговой инженерии
(1970)
Круговая статистика в биологии
(1981)
CircStat: набор инструментов Matlab для круговой статистики
2 J.90 Stat. ПО(2009)
Вариации моделей жизненного цикла морских черепах: неритические и океанические стадии развития
Популяционная генетика и филогеография морских черепах
Мол. Экол.
(2007)
Морские черепахи: естественная история и охрана
(1972)
Прибрежные процессы с инженерными приложениями
(2001)
 Fisher et al.
Fisher et al. Простая функция для множественной регрессии полных подмножеств в экологии с R
Ecol. Эвол.
(2018)
Уровень хищничества рыбами детенышей зеленой черепахи (
Chelonia mydas )Коралловые рифы
(1994)
Генерация лабораторных волн
Проверка чувств: рыбы выбирают новые места обитания, реагируя на многочисленные сигналы
Экология
(2012)
Темы циклической статистики
(2001)
Как личинки лангуста находят берег?
NZ J. Mar. Freshw. Рез.
(2005)
From individual dispersal to species ranges: perspectives for a changing world
Science
(2006)
 J. Limpus
J. LimpusThe flatback turtle, Chelonia depressa Garman in Southeast Queensland, Australia
Herpetologica
( 1971)
Нахождение морских черепах в океане: важность низкого возвышения горизонта в качестве ориентира для ориентации
Поведение
(2013)
Воздействие искусственного освещения в ночное время на морские экосистемы — обзор 2 9036, 2 9036, 9036, 9036, 9036, 2 9036,
,
Биология глобальных изменений
Реакция вылупившихся черепах на воздействие светодиодов в море
2022, Морские и пресноводные исследования
Исследовательская статья
Игра «Чужой против хищника» в прятки: как сложность среды обитания влияет на выживаемость хозяина, опосредованного паразитами
Журнал экспериментальной морской биологии и экологии, том 535, 2021 г., статья 151488 их хозяев, влияя на то, как они реагируют на биотические и абиотические факторы в своих сообществах.
Некоторые паразиты особенно опасны, вызывая значительные репродуктивные и поведенческие изменения у хозяев по сравнению с неинфицированными сородичами. В восточной части Северной Америки неместный паразитический ракушка, Loxothylacus panopaei (Gissler, 1884) оказывает серьезное репродуктивное воздействие на местных панопеидных крабов-хозяев, что приводит к кастрации. Кроме того, было задокументировано, что паразит вызывает значительные поведенческие изменения, такие как снижение активности грязевого краба, влияние на отношения хищник-жертва и усиление скрытого поведения инфицированных крабов-хозяев. Тем не менее многое еще предстоит понять относительно способности хозяина избегать хищников в неоднородной среде. Мы исследовали влияние L. panopaei паразитирование на грязевом крабе (плоскоспинном грязевом крабе, Eurypanopeus depressus (Smith, 1869)) выживание в простых и сложных местообитаниях. В лабораторных экспериментах с мезокосмом неоднородность среды обитания регулировалась с помощью устриц и гравия, а давление хищников исходило от двух обычных хищных крабов в эстуариях юго-восточной Атлантики: атлантического грязевого краба Panopeus herbstii (H. Milne Edwards 1884) и каменного краба . Menippe mercenaria (Сэй, 1818 г.). Мы нашли L. panopaei -инфицированные крабы E. depressus имели самые низкие показатели выживаемости в простой среде обитания из гравия, в то время как крабы с самой высокой выживаемостью были неинфицированными крабами в сложной среде обитания устриц. Кроме того, мы обнаружили различия между неинфицированными и инфицированными крабами в использовании среды обитания (устрицы по сравнению с гравием) в сложной среде обитания. Это исследование демонстрирует сложность динамики хищник-жертва перед лицом разрушительного паразитизма и глобальных изменений, включая интродукцию видов в новые сообщества, где они могут оказать сильное влияние на популяции хозяев и взаимодействие сообществ.Исследовательская статья
Компромиссы между качеством и плодовитостью личинок связаны с материнской средой морского окуня (
Helicolenus percoides, Sebastidae)Journal of Experimental Marine Biology and Ecology, Volume 537, 2021, Article 15152
Пространственные различия в демографии и питании могут привести к различиям в репродуктивной способности и качестве личинок среди морских субпопуляций.
Мы исследовали, как локальные различия в материнском питании влияли на качество и количество личинок морского окуня (9).0111 Helicolenus percoides , Sebastidae), долгоживущий живородящий вид, широко распространенный в Новой Зеландии, с целью определения того, как локальные различия в материнском обеспечении влияют на качество и количество личинок. Сравнивали субпопуляции из Deep Cove в пределах Фьордленда на западном побережье и шельфа Отаго на восточном побережье Южного острова Новой Зеландии. Мы использовали анализ жирных кислот в тканях печени и гонад взрослых, чтобы исследовать потенциальные пространственные различия в питании матерей. Анализ жирных кислот личинок в день отела, длину личиночной хорды и объем масляных глобул в день отела, изменение длины когорт личинок и изменение среднего размера масляных глобул использовали в качестве показателей для установления различий в качестве личинок. между субпопуляциями шельфа Deep Cove и Otago. Наши результаты продемонстрировали компромисс между плодовитостью и размером масляных шариков, где морской окунь с шельфа Отаго был более плодовитым, но снабжал своих личинок меньшими масляными шариками, чем самки из Дип-Коув. Мы наблюдали более быстрое изменение длины и более медленное изменение размера масляного шарика среди когорт личинок, произведенных самками из Deep Cove. Материнские концентрации ключевых жирных кислот докозагексаеновой кислоты (ДГК) и арахидоновой кислоты (АРК) были выше, а соотношение эйкозапентаеновой кислоты (ЭПК)/АРК было ниже среди личинок, произведенных самками из Дип-Коув. Результаты показали, что потенциальные визуальные и осморегуляторные преимущества, связанные с этими жирными кислотами, передаются потомству, что необходимо для выживания в условиях низкой освещенности и низкой продуктивности внутренних фьордов, характеризующихся большим потреблением пресной воды. Результаты настоящего исследования подчеркивают важную связь между материнской средой и качеством и приспособленностью личинок рыб.Исследовательская статья
Галогенсодержащее соединение, выделяемое морскими бактериями, останавливает развитие личинок морских ежей
Журнал экспериментальной морской биологии и экологии, том 538, 2021 г.
, статья 151540Морские бактерии повсеместно распространены, но их экологические функции не выполняются полностью охарактеризован. Распространенный по всему миру образующий биопленку род Pseudoalteromonas производит множество потенциально биоактивных галогенированных органических продуктов. Было обнаружено, что одно такое секретируемое соединение, 2,3,4,5-тетрабромпиррол (ТБФ), является биоцидным для нескольких таксонов фитопланктона, одновременно стимулируя оседание и метаморфоз кораллов в наномолярных дозах. Здесь мы проверили, влияет ли присутствие ТБФ на раннее развитие морских ежей, которые не находятся в прямом контакте с бентосной микробной пленкой. 9Эмбрионы 0111 Lytechinus variegatus подвергались воздействию различных концентраций ТБФ в течение различных периодов времени в течение первых 48 часов после оплодотворения. Такие низкие концентрации, как 500 нМ, заметно снижали выживаемость личинок и замедляли развитие. Эти вредные эффекты становились более выраженными по мере увеличения концентрации и продолжительности воздействия ТБФ.
Нарушения в развитии при воздействии ТБФ оказались обратимыми при низких концентрациях воздействия (< 500 нМ – 1000 нМ) при ограниченной продолжительности воздействия (< 1–4 ч). Предварительное иммунофлуоресцентное окрашивание выявило дефекты веретена деления у делящихся эмбрионов при воздействии ТБФ в высоких концентрациях, что могло способствовать гибели и нарушению роста. В то время как ТБФ и другие подобные ему бактериальные соединения могут служить сигналами для заселения кораллов, их цитотоксичность по отношению к одноклеточным водорослям и личиночным ежам может указывать на опасность бентоса для некоторых развивающихся эмбрионов беспозвоночных. Если это правда, то такие соединения вместе с таксонами бактерий, которые их производят, вероятно, играют недооцененную роль в экологии и, возможно, в эволюции планктонного развития макробеспозвоночных.Исследовательская статья
Местная среда формирует адаптацию
Phaeocystis antarctica к возмущениям солености: доказательства физиологической устойчивостиЖурнал экспериментальной морской биологии и экологии, том 538, 2021 г.
, статья 1510377 The Southern Ocean) является хрупкой экосистемой, судя по изменениям времени наступления и отступления ее ледяного покрова. В ЮК Антарктическое циркумполярное течение (ACC) и прибрежные круговороты (море Уэдделла и море Росса) обеспечивают локальные условия с различными характеристиками температуры и солености, связанными с различной историей морского льда. Phaeocystis antarctica является примнезиофитом, часто доминирующим в цветении полярного фитопланктона в (SO), и является там ключевым видом, поскольку его численность может оказывать негативное влияние на более высокие трофические уровни и может влиять на газообмен воздух/море, участвующий в производстве ДМСП. Таким образом, его способность выживать в ответ на возмущения окружающей среды может быть связана с его генетическим разнообразием в его популяциях, когда они перемещаются по SO. Здесь мы применяем повышенную (70 PSU) и пониженную (18 PSU) обработку соленостью к пяти генетически различным Штаммы P.
antarctica , выделенные из трех различных водных масс, для проверки того, что более важно: генетическое сходство или физические характеристики водной массы в определении реакции на изменения солености, например, возникающие при включении в рассоловые каналы морского льда и/или его последующую талую воду в тех водных массах, которые имеют годовой ледовый покров. Штаммы, которые были географически близки (выделены из одной и той же водной массы), но генетически различались (около 30% сходны и принадлежали к разным генофондам, судя по анализу микросателлитного (МС) и амплифицированного фрагмент-связанного полиморфизма (AFLP), одинаково реагировали на более высокие и более низкие солености, тогда как генетически близкие штаммы (около 95% идентичных или из одного и того же генофонда), происходящих из разных водных масс и, следовательно, разных условий среды, реагировали по-разному. Продукция диметилсульфониопропионата (DMSP) в ответ на эти изменения солености существенно не различалась между любыми штаммами/обработками. Учитывая наличие очень сходных генотипов на свободных ото льда, а также на сезонно покрытых льдом участках отбора проб, наблюдаемые фенотипические различия, скорее всего, являются результатом быстрой локальной адаптации между обоими типами местообитаний или гибридизацией изолятов из двух генофондов с результирующим генотипом для солеустойчивость является той, из которой произошел изолят, что позволяет предположить, что избирательный эффект сезонного присутствия или круглогодичного отсутствия морского льда преобладает над потоком генов между этими средами обитания, чтобы адаптировать физиологию клеток к окружающей среде за относительно короткий период времени.Исследовательская статья
Сиртуин-зависимое восстановление после воздушного теплового шока: влияние пищевого рациона, термической истории и ингибирования сиртуином на скорость клиренса и активность раскрытия створки калифорнийской мидии,
Mytilus californianus (Conrad)Журнал экспериментальной морской биологии и экологии, том 536, 2021 г.
, статья 151510Приливные мидии регулярно испытывают сезонные колебания в доступности пищи и тепловой стресс в течение приливного цикла, однако мало что известно о биохимических механизмах, связывающих пищевой рацион и температурный режим. толерантность, несмотря на доказательства того, что пища повышает толерантность к стрессу на уровне организма. Мы предположили, что сиртуины (пищевые белковые регуляторы клеточного гомеостаза), которые могут регулировать клеточную стрессовую реакцию митилид во время теплового стресса, также влияют на работоспособность организма во время восстановления после теплового стресса. В рамках более широкого исследования мы исследовали влияние акклиматизационного рациона питания, температуры всплытия (воздуха) и ингибиторов сиртуина на пищевое поведение калифорнийской мидии, Mytilus californianus, во время восстановления после острого воздушного теплового шока. Мидии акклиматизировались в течение 3 недель. к низкому (0,25% водорослей·г сухого веса мидий -1 ) или высокому (1,5% водорослей·г сухого веса мидии -1 ) рациону водорослей и низкому (20 °C) или высокому (30 °C) рациону температуры всплытия в резервуарах-симуляторах приливов, способных поддерживать стандартные циркадные и циркадиальные ритмы.
После акклиматизации мидии подвергались воздействию двух ингибиторов сиртуина (никотинамид и сурамин) или не подвергались воздействию ингибиторов в течение одного цикла прилива. Во время последующего цикла отлива все мидии подверглись острому воздушному тепловому шоку (33 °C), а затем им позволили восстановиться в соответствующих условиях акклиматизации. Каждые 12 часов в течение 48 часов до ингибирования сиртуинов и 48 часов после острого теплового шока (в соответствующие периоды приливов) мы регистрировали скорость клиренса и активность открытия клапана. Мидии, акклиматизированные к низкому рациону питания, значительно снижали скорость выведения при тепловом шоке и ингибировании сиртуинов, но не только при тепловом шоке, по сравнению со значениями до обработки, независимо от температуры всплытия. У мидий, акклиматизированных к низкому пищевому рациону и высокой температуре всплытия, наблюдалось значительное снижение активности щели створки после острого теплового шока после ингибирования сиртуина, но не в других группах акклиматизации. Неожиданно тепловой шок сам по себе снизил скорость выведения мидий, акклиматизированных только к высокой температуре пищи и высокой температуре всплытия, без какого-либо дополнительного эффекта теплового шока и ингибирования сиртуинов. В совокупности эти данные свидетельствуют о важной роли сиртуинов на уровне организма во время восстановления после теплового шока после адаптации к низкому пищевому рациону.Исследовательская статья
Изменения условий окружающей среды в пределах растительного покрова и структуры популяции крапчатого гребешка, связанные с локальной потерей надземного покрова морских водорослей
Journal of Experimental Marine Biology and Ecology, Volume 534, 2021, Article 151486
Трехмерная структура морских водорослей способствует разнообразию сообществ, предоставляя убежище от хищников, возможности поиска пищи и смягчения физического стресса. Влияние структурной сложности морских водорослей на связанные сообщества было связано с биомассой и плотностью побегов морских водорослей, часто с использованием экспериментов с мезокосмом, которые не могут учитывать естественные физические условия.
Здесь мы манипулируем травяным покровом в поле, чтобы определить краткосрочные и локальные последствия потери Zostera marina (морская трава) на физические условия под пологом, а также на распределение размеров и смертность обыкновенного двустворчатого моллюска, морского гребешка Argopecten circleis . По сравнению с нетронутой взморником (100% покрытие), на участках с 50% покрытием или без покрытия (100% обнаженных отложений) наблюдается значительное увеличение среднего дневного освещения и температуры. Изменения относительной смертности морского гребешка были связаны с сокращением площади морского морского гребешка, а уменьшение размера морского гребешка было связано с изменением освещенности и температуры после обрезки морского гребешка. Эти результаты подчеркивают важность покрова морских водорослей для регулирования условий окружающей среды под пологом и потенциальной модуляции биологических взаимодействий связанных с ними беспозвоночных.
 Некоторые паразиты особенно опасны, вызывая значительные репродуктивные и поведенческие изменения у хозяев по сравнению с неинфицированными сородичами. В восточной части Северной Америки неместный паразитический ракушка, Loxothylacus panopaei (Gissler, 1884) оказывает серьезное репродуктивное воздействие на местных панопеидных крабов-хозяев, что приводит к кастрации. Кроме того, было задокументировано, что паразит вызывает значительные поведенческие изменения, такие как снижение активности грязевого краба, влияние на отношения хищник-жертва и усиление скрытого поведения инфицированных крабов-хозяев. Тем не менее многое еще предстоит понять относительно способности хозяина избегать хищников в неоднородной среде. Мы исследовали влияние L. panopaei паразитирование на грязевом крабе (плоскоспинном грязевом крабе, Eurypanopeus depressus (Smith, 1869)) выживание в простых и сложных местообитаниях. В лабораторных экспериментах с мезокосмом неоднородность среды обитания регулировалась с помощью устриц и гравия, а давление хищников исходило от двух обычных хищных крабов в эстуариях юго-восточной Атлантики: атлантического грязевого краба Panopeus herbstii (H.
Некоторые паразиты особенно опасны, вызывая значительные репродуктивные и поведенческие изменения у хозяев по сравнению с неинфицированными сородичами. В восточной части Северной Америки неместный паразитический ракушка, Loxothylacus panopaei (Gissler, 1884) оказывает серьезное репродуктивное воздействие на местных панопеидных крабов-хозяев, что приводит к кастрации. Кроме того, было задокументировано, что паразит вызывает значительные поведенческие изменения, такие как снижение активности грязевого краба, влияние на отношения хищник-жертва и усиление скрытого поведения инфицированных крабов-хозяев. Тем не менее многое еще предстоит понять относительно способности хозяина избегать хищников в неоднородной среде. Мы исследовали влияние L. panopaei паразитирование на грязевом крабе (плоскоспинном грязевом крабе, Eurypanopeus depressus (Smith, 1869)) выживание в простых и сложных местообитаниях. В лабораторных экспериментах с мезокосмом неоднородность среды обитания регулировалась с помощью устриц и гравия, а давление хищников исходило от двух обычных хищных крабов в эстуариях юго-восточной Атлантики: атлантического грязевого краба Panopeus herbstii (H. Milne Edwards 1884) и каменного краба . Menippe mercenaria (Сэй, 1818 г.). Мы нашли L. panopaei -инфицированные крабы E. depressus имели самые низкие показатели выживаемости в простой среде обитания из гравия, в то время как крабы с самой высокой выживаемостью были неинфицированными крабами в сложной среде обитания устриц. Кроме того, мы обнаружили различия между неинфицированными и инфицированными крабами в использовании среды обитания (устрицы по сравнению с гравием) в сложной среде обитания. Это исследование демонстрирует сложность динамики хищник-жертва перед лицом разрушительного паразитизма и глобальных изменений, включая интродукцию видов в новые сообщества, где они могут оказать сильное влияние на популяции хозяев и взаимодействие сообществ.
Milne Edwards 1884) и каменного краба . Menippe mercenaria (Сэй, 1818 г.). Мы нашли L. panopaei -инфицированные крабы E. depressus имели самые низкие показатели выживаемости в простой среде обитания из гравия, в то время как крабы с самой высокой выживаемостью были неинфицированными крабами в сложной среде обитания устриц. Кроме того, мы обнаружили различия между неинфицированными и инфицированными крабами в использовании среды обитания (устрицы по сравнению с гравием) в сложной среде обитания. Это исследование демонстрирует сложность динамики хищник-жертва перед лицом разрушительного паразитизма и глобальных изменений, включая интродукцию видов в новые сообщества, где они могут оказать сильное влияние на популяции хозяев и взаимодействие сообществ. Мы исследовали, как локальные различия в материнском питании влияли на качество и количество личинок морского окуня (9).0111 Helicolenus percoides , Sebastidae), долгоживущий живородящий вид, широко распространенный в Новой Зеландии, с целью определения того, как локальные различия в материнском обеспечении влияют на качество и количество личинок. Сравнивали субпопуляции из Deep Cove в пределах Фьордленда на западном побережье и шельфа Отаго на восточном побережье Южного острова Новой Зеландии. Мы использовали анализ жирных кислот в тканях печени и гонад взрослых, чтобы исследовать потенциальные пространственные различия в питании матерей. Анализ жирных кислот личинок в день отела, длину личиночной хорды и объем масляных глобул в день отела, изменение длины когорт личинок и изменение среднего размера масляных глобул использовали в качестве показателей для установления различий в качестве личинок. между субпопуляциями шельфа Deep Cove и Otago. Наши результаты продемонстрировали компромисс между плодовитостью и размером масляных шариков, где морской окунь с шельфа Отаго был более плодовитым, но снабжал своих личинок меньшими масляными шариками, чем самки из Дип-Коув.
Мы исследовали, как локальные различия в материнском питании влияли на качество и количество личинок морского окуня (9).0111 Helicolenus percoides , Sebastidae), долгоживущий живородящий вид, широко распространенный в Новой Зеландии, с целью определения того, как локальные различия в материнском обеспечении влияют на качество и количество личинок. Сравнивали субпопуляции из Deep Cove в пределах Фьордленда на западном побережье и шельфа Отаго на восточном побережье Южного острова Новой Зеландии. Мы использовали анализ жирных кислот в тканях печени и гонад взрослых, чтобы исследовать потенциальные пространственные различия в питании матерей. Анализ жирных кислот личинок в день отела, длину личиночной хорды и объем масляных глобул в день отела, изменение длины когорт личинок и изменение среднего размера масляных глобул использовали в качестве показателей для установления различий в качестве личинок. между субпопуляциями шельфа Deep Cove и Otago. Наши результаты продемонстрировали компромисс между плодовитостью и размером масляных шариков, где морской окунь с шельфа Отаго был более плодовитым, но снабжал своих личинок меньшими масляными шариками, чем самки из Дип-Коув. Мы наблюдали более быстрое изменение длины и более медленное изменение размера масляного шарика среди когорт личинок, произведенных самками из Deep Cove. Материнские концентрации ключевых жирных кислот докозагексаеновой кислоты (ДГК) и арахидоновой кислоты (АРК) были выше, а соотношение эйкозапентаеновой кислоты (ЭПК)/АРК было ниже среди личинок, произведенных самками из Дип-Коув. Результаты показали, что потенциальные визуальные и осморегуляторные преимущества, связанные с этими жирными кислотами, передаются потомству, что необходимо для выживания в условиях низкой освещенности и низкой продуктивности внутренних фьордов, характеризующихся большим потреблением пресной воды. Результаты настоящего исследования подчеркивают важную связь между материнской средой и качеством и приспособленностью личинок рыб.
Мы наблюдали более быстрое изменение длины и более медленное изменение размера масляного шарика среди когорт личинок, произведенных самками из Deep Cove. Материнские концентрации ключевых жирных кислот докозагексаеновой кислоты (ДГК) и арахидоновой кислоты (АРК) были выше, а соотношение эйкозапентаеновой кислоты (ЭПК)/АРК было ниже среди личинок, произведенных самками из Дип-Коув. Результаты показали, что потенциальные визуальные и осморегуляторные преимущества, связанные с этими жирными кислотами, передаются потомству, что необходимо для выживания в условиях низкой освещенности и низкой продуктивности внутренних фьордов, характеризующихся большим потреблением пресной воды. Результаты настоящего исследования подчеркивают важную связь между материнской средой и качеством и приспособленностью личинок рыб. , статья 151540
, статья 151540 Нарушения в развитии при воздействии ТБФ оказались обратимыми при низких концентрациях воздействия (< 500 нМ – 1000 нМ) при ограниченной продолжительности воздействия (< 1–4 ч). Предварительное иммунофлуоресцентное окрашивание выявило дефекты веретена деления у делящихся эмбрионов при воздействии ТБФ в высоких концентрациях, что могло способствовать гибели и нарушению роста. В то время как ТБФ и другие подобные ему бактериальные соединения могут служить сигналами для заселения кораллов, их цитотоксичность по отношению к одноклеточным водорослям и личиночным ежам может указывать на опасность бентоса для некоторых развивающихся эмбрионов беспозвоночных. Если это правда, то такие соединения вместе с таксонами бактерий, которые их производят, вероятно, играют недооцененную роль в экологии и, возможно, в эволюции планктонного развития макробеспозвоночных.
Нарушения в развитии при воздействии ТБФ оказались обратимыми при низких концентрациях воздействия (< 500 нМ – 1000 нМ) при ограниченной продолжительности воздействия (< 1–4 ч). Предварительное иммунофлуоресцентное окрашивание выявило дефекты веретена деления у делящихся эмбрионов при воздействии ТБФ в высоких концентрациях, что могло способствовать гибели и нарушению роста. В то время как ТБФ и другие подобные ему бактериальные соединения могут служить сигналами для заселения кораллов, их цитотоксичность по отношению к одноклеточным водорослям и личиночным ежам может указывать на опасность бентоса для некоторых развивающихся эмбрионов беспозвоночных. Если это правда, то такие соединения вместе с таксонами бактерий, которые их производят, вероятно, играют недооцененную роль в экологии и, возможно, в эволюции планктонного развития макробеспозвоночных.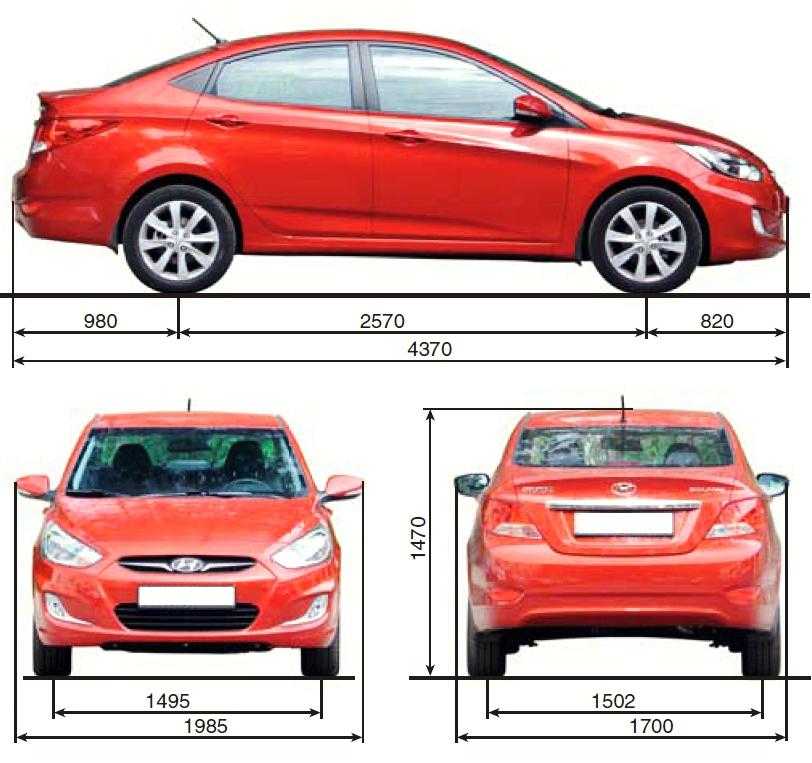 , статья 151037
, статья 151037 antarctica , выделенные из трех различных водных масс, для проверки того, что более важно: генетическое сходство или физические характеристики водной массы в определении реакции на изменения солености, например, возникающие при включении в рассоловые каналы морского льда и/или его последующую талую воду в тех водных массах, которые имеют годовой ледовый покров. Штаммы, которые были географически близки (выделены из одной и той же водной массы), но генетически различались (около 30% сходны и принадлежали к разным генофондам, судя по анализу микросателлитного (МС) и амплифицированного фрагмент-связанного полиморфизма (AFLP), одинаково реагировали на более высокие и более низкие солености, тогда как генетически близкие штаммы (около 95% идентичных или из одного и того же генофонда), происходящих из разных водных масс и, следовательно, разных условий среды, реагировали по-разному. Продукция диметилсульфониопропионата (DMSP) в ответ на эти изменения солености существенно не различалась между любыми штаммами/обработками.
antarctica , выделенные из трех различных водных масс, для проверки того, что более важно: генетическое сходство или физические характеристики водной массы в определении реакции на изменения солености, например, возникающие при включении в рассоловые каналы морского льда и/или его последующую талую воду в тех водных массах, которые имеют годовой ледовый покров. Штаммы, которые были географически близки (выделены из одной и той же водной массы), но генетически различались (около 30% сходны и принадлежали к разным генофондам, судя по анализу микросателлитного (МС) и амплифицированного фрагмент-связанного полиморфизма (AFLP), одинаково реагировали на более высокие и более низкие солености, тогда как генетически близкие штаммы (около 95% идентичных или из одного и того же генофонда), происходящих из разных водных масс и, следовательно, разных условий среды, реагировали по-разному. Продукция диметилсульфониопропионата (DMSP) в ответ на эти изменения солености существенно не различалась между любыми штаммами/обработками. Учитывая наличие очень сходных генотипов на свободных ото льда, а также на сезонно покрытых льдом участках отбора проб, наблюдаемые фенотипические различия, скорее всего, являются результатом быстрой локальной адаптации между обоими типами местообитаний или гибридизацией изолятов из двух генофондов с результирующим генотипом для солеустойчивость является той, из которой произошел изолят, что позволяет предположить, что избирательный эффект сезонного присутствия или круглогодичного отсутствия морского льда преобладает над потоком генов между этими средами обитания, чтобы адаптировать физиологию клеток к окружающей среде за относительно короткий период времени.
Учитывая наличие очень сходных генотипов на свободных ото льда, а также на сезонно покрытых льдом участках отбора проб, наблюдаемые фенотипические различия, скорее всего, являются результатом быстрой локальной адаптации между обоими типами местообитаний или гибридизацией изолятов из двух генофондов с результирующим генотипом для солеустойчивость является той, из которой произошел изолят, что позволяет предположить, что избирательный эффект сезонного присутствия или круглогодичного отсутствия морского льда преобладает над потоком генов между этими средами обитания, чтобы адаптировать физиологию клеток к окружающей среде за относительно короткий период времени. , статья 151510
, статья 151510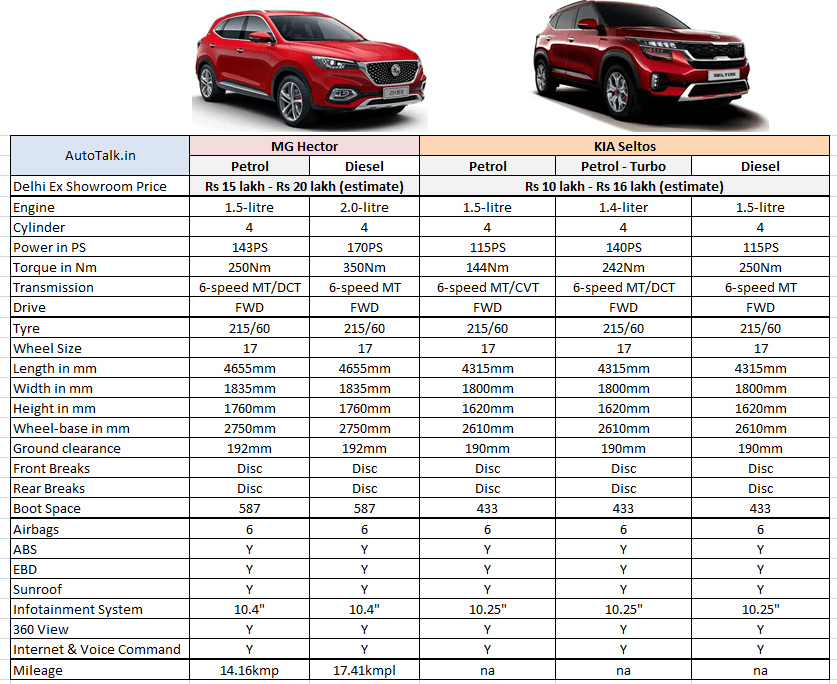 После акклиматизации мидии подвергались воздействию двух ингибиторов сиртуина (никотинамид и сурамин) или не подвергались воздействию ингибиторов в течение одного цикла прилива. Во время последующего цикла отлива все мидии подверглись острому воздушному тепловому шоку (33 °C), а затем им позволили восстановиться в соответствующих условиях акклиматизации. Каждые 12 часов в течение 48 часов до ингибирования сиртуинов и 48 часов после острого теплового шока (в соответствующие периоды приливов) мы регистрировали скорость клиренса и активность открытия клапана. Мидии, акклиматизированные к низкому рациону питания, значительно снижали скорость выведения при тепловом шоке и ингибировании сиртуинов, но не только при тепловом шоке, по сравнению со значениями до обработки, независимо от температуры всплытия. У мидий, акклиматизированных к низкому пищевому рациону и высокой температуре всплытия, наблюдалось значительное снижение активности щели створки после острого теплового шока после ингибирования сиртуина, но не в других группах акклиматизации.
После акклиматизации мидии подвергались воздействию двух ингибиторов сиртуина (никотинамид и сурамин) или не подвергались воздействию ингибиторов в течение одного цикла прилива. Во время последующего цикла отлива все мидии подверглись острому воздушному тепловому шоку (33 °C), а затем им позволили восстановиться в соответствующих условиях акклиматизации. Каждые 12 часов в течение 48 часов до ингибирования сиртуинов и 48 часов после острого теплового шока (в соответствующие периоды приливов) мы регистрировали скорость клиренса и активность открытия клапана. Мидии, акклиматизированные к низкому рациону питания, значительно снижали скорость выведения при тепловом шоке и ингибировании сиртуинов, но не только при тепловом шоке, по сравнению со значениями до обработки, независимо от температуры всплытия. У мидий, акклиматизированных к низкому пищевому рациону и высокой температуре всплытия, наблюдалось значительное снижение активности щели створки после острого теплового шока после ингибирования сиртуина, но не в других группах акклиматизации. Неожиданно тепловой шок сам по себе снизил скорость выведения мидий, акклиматизированных только к высокой температуре пищи и высокой температуре всплытия, без какого-либо дополнительного эффекта теплового шока и ингибирования сиртуинов. В совокупности эти данные свидетельствуют о важной роли сиртуинов на уровне организма во время восстановления после теплового шока после адаптации к низкому пищевому рациону.
Неожиданно тепловой шок сам по себе снизил скорость выведения мидий, акклиматизированных только к высокой температуре пищи и высокой температуре всплытия, без какого-либо дополнительного эффекта теплового шока и ингибирования сиртуинов. В совокупности эти данные свидетельствуют о важной роли сиртуинов на уровне организма во время восстановления после теплового шока после адаптации к низкому пищевому рациону. Здесь мы манипулируем травяным покровом в поле, чтобы определить краткосрочные и локальные последствия потери Zostera marina (морская трава) на физические условия под пологом, а также на распределение размеров и смертность обыкновенного двустворчатого моллюска, морского гребешка Argopecten circleis . По сравнению с нетронутой взморником (100% покрытие), на участках с 50% покрытием или без покрытия (100% обнаженных отложений) наблюдается значительное увеличение среднего дневного освещения и температуры. Изменения относительной смертности морского гребешка были связаны с сокращением площади морского морского гребешка, а уменьшение размера морского гребешка было связано с изменением освещенности и температуры после обрезки морского гребешка. Эти результаты подчеркивают важность покрова морских водорослей для регулирования условий окружающей среды под пологом и потенциальной модуляции биологических взаимодействий связанных с ними беспозвоночных.
Здесь мы манипулируем травяным покровом в поле, чтобы определить краткосрочные и локальные последствия потери Zostera marina (морская трава) на физические условия под пологом, а также на распределение размеров и смертность обыкновенного двустворчатого моллюска, морского гребешка Argopecten circleis . По сравнению с нетронутой взморником (100% покрытие), на участках с 50% покрытием или без покрытия (100% обнаженных отложений) наблюдается значительное увеличение среднего дневного освещения и температуры. Изменения относительной смертности морского гребешка были связаны с сокращением площади морского морского гребешка, а уменьшение размера морского гребешка было связано с изменением освещенности и температуры после обрезки морского гребешка. Эти результаты подчеркивают важность покрова морских водорослей для регулирования условий окружающей среды под пологом и потенциальной модуляции биологических взаимодействий связанных с ними беспозвоночных. Crown Copyright © 2020 Опубликовано Elsevier B. V. Все права защищены.
V. Все права защищены.
Научные заметки 2001: Сообразительный морской лев
Рио, калифорнийский морской лев, плавает вокруг
тренировочный бассейн, как встревоженный спортсмен перед гонкой. Она готовится
за ее главную роль в эксперименте, предназначенном для проверки ее способности
считать. Стремясь начать, она выражает свое нетерпение звонком, который звучит
как нечто среднее между львиным рыком и кваканьем жабы
Рио может нервничать, когда собирается провести свой эксперимент, отмечает Шеннон Спилман, тренер, наблюдающий за лабораторией. Немного нервничает, делает глубокий вдох, надевает гарнитуру с микрофоном, учится свои записи на мгновение, кладет руки на клавиатуру и проверяет Деятельность Рио на видеоэкране. Она похожа на авиадиспетчера. «За этим нужно следить, — говорит она.
Готовы? Спиллман спрашивает студентов-добровольцев снаружи в
бассейн. «Хорошо, пошли». И шоу начинается.
И шоу начинается.
Один из волонтеров звонит в Рио. Она выпрыгивает из воды и грациозно
скользит к остановке перед деревянным экраном у бассейна, который выглядит немного
как декорации телевизионного игрового шоу. Она кладет голову на подставку лицом к
три окна. Центральное окно открывается, чтобы показать квадратную карту, немного
высотой более фута, с черно-белым изображением корабля.
якорь на нем. Рио несколько секунд изучает картинку, прежде чем
окна открываются, обнажая изображения кактуса слева и воющего
волк справа. Рио быстро оглядывается на символы и
потом ждет. Спиллман набирает ноту на клавиатуре, чтобы подать сигнал Рио, который тут же
прикасается носом к изображению волка. Спилман берет более высокую ноту
означает правильный ответ, а Рио ловит рыбу, которую бросает волонтер.
из-за экрана. Видимо, все согласны с тем, что волк как-то
связанные с якорем.
Этот загадочный эксперимент на самом деле является демонстрацией способности Рио
думать.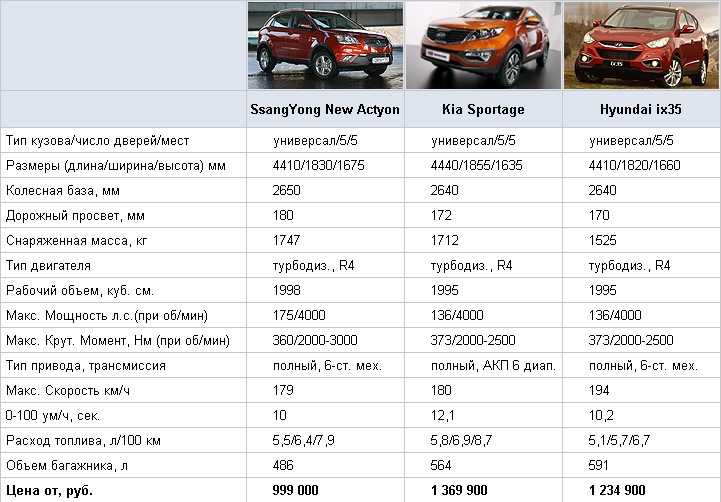 Она удивила исследователей логическими подвигами без животных
кроме того, что люди сделали в лаборатории. Рио и ее когорта,
Рокки, продемонстрировали способность делать логические выводы,
ранее считалось, что для этого требуется разговорный язык. Теперь кажется, что язык требует
логика, а не наоборот. Это исключительно важное исследование приходит
из лаборатории Рональда Шустермана в Калифорнийском университете в Санта
Круз, который годами кропотливо дрессировал морских львов.
Она удивила исследователей логическими подвигами без животных
кроме того, что люди сделали в лаборатории. Рио и ее когорта,
Рокки, продемонстрировали способность делать логические выводы,
ранее считалось, что для этого требуется разговорный язык. Теперь кажется, что язык требует
логика, а не наоборот. Это исключительно важное исследование приходит
из лаборатории Рональда Шустермана в Калифорнийском университете в Санта
Круз, который годами кропотливо дрессировал морских львов.
Шустерман считает, что эти способности могли развиться, чтобы помогать животным решать сложные социальные проблемы в дикой природе, например узнавать друзей и врагов, используя множество различных сигналов, таких как зрение, запах и звук. «Это действительно расширяет наши представления о том, как животное воспринимает свой мир», он сказал.
Интерес Шустермана к разуму животных начался в 1960 году в Йерксе.
Лаборатории биологии приматов в Ориндж-Парке, Флорида, где он исследовал
когнитивное и социальное поведение шимпанзе, гиббонов и обезьян. В 19В 63 году он перешел в Стэнфордский научно-исследовательский институт, чтобы создать первую
лаборатории в Северной Америке, посвященной поведению и физиологии
тюлени и морские львы. Шустерман и его морские львы провели несколько лет в
Калифорнийский государственный университет в Хейворде, прежде чем они переехали в университет
Калифорнийской лаборатории Long Marine в Санта-Круз, где они
остаться сегодня. «Везде, где я мог бы привести своих морских львов,
Вот куда я пошел, — сказал он.
В 19В 63 году он перешел в Стэнфордский научно-исследовательский институт, чтобы создать первую
лаборатории в Северной Америке, посвященной поведению и физиологии
тюлени и морские львы. Шустерман и его морские львы провели несколько лет в
Калифорнийский государственный университет в Хейворде, прежде чем они переехали в университет
Калифорнийской лаборатории Long Marine в Санта-Круз, где они
остаться сегодня. «Везде, где я мог бы привести своих морских львов,
Вот куда я пошел, — сказал он.
Рокки и Рио — мировые звезды в научном сообществе и также в глазах общественности. Они были представлены в журнале Time и USA Today и появились в телешоу PBS под названием «Животные». Эйнштейны.
Призовый ученик Шустермана Рио родился в неволе 15 лет назад.
и воспитан суррогатной матерью-человеком. Она энергичная, игривая ученица
и, кажется, наслаждается своей ролью в исследовании. Ее мотивация и внимание
сделать ее идеальным предметом.
Рио может быть звездой шоу, но Рокки — настоящий ветеран. Заброшенный в дикой природе щенком, она была с Шустерманом с тех пор, как ее спасли в возраст один. 25 лет Рокки значительно превышают средний возраст 16 лет. продолжительность жизни дикого калифорнийского морского льва. Несмотря на катаракту, препятствующую ее способность проводить эксперименты, она продолжает быть добровольным участником. При весе 150 фунтов она меньше средней самки морского льва, но ее вклад в наше понимание животного разума несет в себе много масса.
Вдохновленный исследованиями афалин в Гавайском университете, Шустерман научила Рокки языку жестов в 10 лет. Она научилась выступать. сложные задачи, включающие несколько знаков, например, прикосновение к маленькому белому мяч ластами или поднесение большого черного конуса к маленькому черному мячу.
Отдельные дельфины и приматы уже овладели знаками
язык, но Рокки тем более примечателен тем, что очищает тот же умственный
препятствие с меньшим мозгом. Ее достижение заставило Шустермана предположить
что он имел дело с очень простым процессом обучения, который мог
корень многих сложных моделей поведения. Поэтому он решил раскрыть основную
механизм.
Ее достижение заставило Шустермана предположить
что он имел дело с очень простым процессом обучения, который мог
корень многих сложных моделей поведения. Поэтому он решил раскрыть основную
механизм.
Поиски Шустермана привели его в область детской психологии. Он
модифицированные методы, используемые для выявления языкового понимания у отсталых детей
и разработали методы для достижения тех же результатов с морскими львами. За
лет он неукоснительно обучал Рокки и Рио навыкам и информации
им нужно добиться успеха.
Морские львы научились сопоставлять множество пар символов путем проб и ошибок.
ошибка, с рыбой в качестве положительного подкрепления. Когда-то у них была твердая хватка
отношений между парами, животным был предоставлен новый выбор
которые не были непосредственно обучены видеть, как они будут реагировать. Например,
они заранее узнали, что изображение замка сочетается с
гольф-клуб, и что гольф-клуб идет с пиццей. Но могли ли они
сделать логическую связь, что пицца идет с замком?
Но могли ли они
сделать логическую связь, что пицца идет с замком?
Для Рио ответ убедительный: да. Она сразу знала ответ, и она ответила правильно в первый раз, когда ей дали тот же тип по выбору с каждой группой символов.
В это было трудно поверить, когда мы получили результаты, Шустерман. сказал о первой демонстрации в Рио. Этот интеллектуальный скачок доказал что Рио могла формировать в уме логические связи, не непосредственно обучены. До успеха Рио-де-Жанейро многие ученые считали что способность устанавливать эти связи, известная как «эквивалентность», было уникальной человеческой характеристикой, невозможной без разговорного языка. Напротив, это может быть важным строительным блоком для языка.
Доктор. Шустерман единственный, кто, я думаю, однозначно
продемонстрировал эквивалентность у нечеловеческих испытуемых, — сказал Мюррей Сидман из
Детский центр Новой Англии в Саутборо, штат Массачусетс. Из
вся работа с нелюдьми, она больше всего напоминает то, что мы видим с человеческими
существ. Исследования человеческих младенцев показали, что эти логические
способности предшествуют способности говорить. Аналогичные исследования с
у умственно отсталых детей также проявлялось логическое мышление при отсутствии
речь.
Из
вся работа с нелюдьми, она больше всего напоминает то, что мы видим с человеческими
существ. Исследования человеческих младенцев показали, что эти логические
способности предшествуют способности говорить. Аналогичные исследования с
у умственно отсталых детей также проявлялось логическое мышление при отсутствии
речь.
Сидман был первым исследователем, продемонстрировавшим эту способность у не говорящего
человек. Он считается отцом теории эквивалентности, которую часто называют
как «эквивалентность Сидмана». В его исследовании 1971 года использовалась эквивалентность
объяснить развитие понимания прочитанного у глубоко отсталого
мальчик-подросток. В этом основополагающем эксперименте три «символа»
должны были соответствовать изображение, печатное слово и произносимое слово. Мальчик
учили реагировать на произносимое слово «стул» выбором
изображение стула или печатное слово C-H-A-I-R. Мальчик был тогда
способен замкнуть логический цикл, отвечая на написанное слово выбором
картинка и наоборот.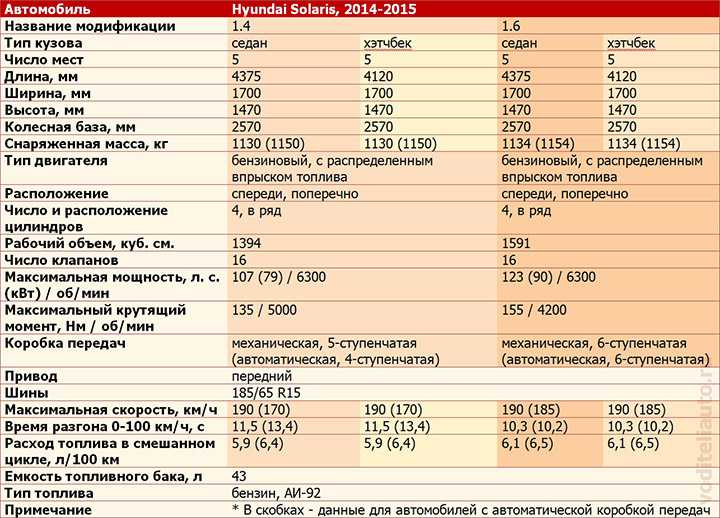
Чтобы иметь язык, люди должны иметь возможность придавать значения слова. Одна из теорий состоит в том, что набор ассоциаций эквивалентности между Вербальное и визуальное представление объекта лежит в основе нашей способности говорить. Человеческие младенцы начинают устанавливать эти связи еще до того, как способен их вербализовать. Как только они начинают говорить, легко смотреть набор ассоциаций растет по мере связывания слова «мама» видом своих матерей или словом «печенье» своими любимая закуска.
«В языке нет ничего волшебного», — сказал Уильям Дьюб. Э.К. Центр умственной отсталости Шрайвера в Уолтеме, Массачусетс, который также проводил работу по эквивалентности с людьми. Одна из вещей в основе нашего языка лежит эта сеть символических отношений. Вы не изучаете все отношения непосредственно, но вы можете вести себя так, как будто все они были непосредственно обучены.
Шустерман считает, что Рио показал нам, что эквивалентность может быть не
шаг в развитии человеческого языка, но и этап в эволюции
языка. Я считаю, что эквивалентность является предшественником языка
в эволюционном смысле, — сказал Шустерман. Морские львы не
развили свой язык. Но они, возможно, заложили фундамент, предлагая
взгляд на таинственную эволюцию человеческого языка.
Я считаю, что эквивалентность является предшественником языка
в эволюционном смысле, — сказал Шустерман. Морские львы не
развили свой язык. Но они, возможно, заложили фундамент, предлагая
взгляд на таинственную эволюцию человеческого языка.
Однако некоторые исследователи, работающие с людьми, ставят под сомнение важность
демонстрации в Рио. Стивен Хейс, психолог из университета
Невады в Рино, скептически относится к модификациям, сделанным Шустерманом.
к методам, используемым с людьми. В отличие от Хейса и некоторых его коллег,
Шустерман использует несколько примеров одной и той же проблемы, чтобы помочь морю
Львы понимают, о чем их просят. Он также использует рыбные награды.
как усиление. Хейс чувствует, что награды и повторение омрачают
результаты, достижения. «Очень сложно понять, функционально ли это
то же самое, что мы видим у людей. Я хочу, чтобы это было сделано именно так,
как мы получаем его у 18-месячных младенцев, сказал он.
Шустерман поддерживает свою технику. Как и в любом эксперименте с животными в лаборатории, где вас интересует ответ животного вопрос, вы должны убедиться, что животное понимает природу вопрос, сказал он. С людьми вы можете говорить с ними и дать им инструкции. С животными вы можете дать только много Примеры. Как только вы предоставите эти примеры, вы сможете настроить тестирование. процедура. Это то, что мы сделали».
Воодушевленные своим успехом, Шустерман и его аспиранты продолжают
чтобы проверить пределы психики морского льва. Коллин Кастак, выпускница
студент, работающий с Шустерманом почти 10 лет,
был вовлечен в работу по эквивалентности практически с самого начала. В течение
в то время она неустанно работала с Рокки и Рио над расширением
начальная демонстрация эквивалентности. Ее последняя работа представляет
удивительный и важный шаг: доказать, что морские львы могут организовать
символы на функциональные категории.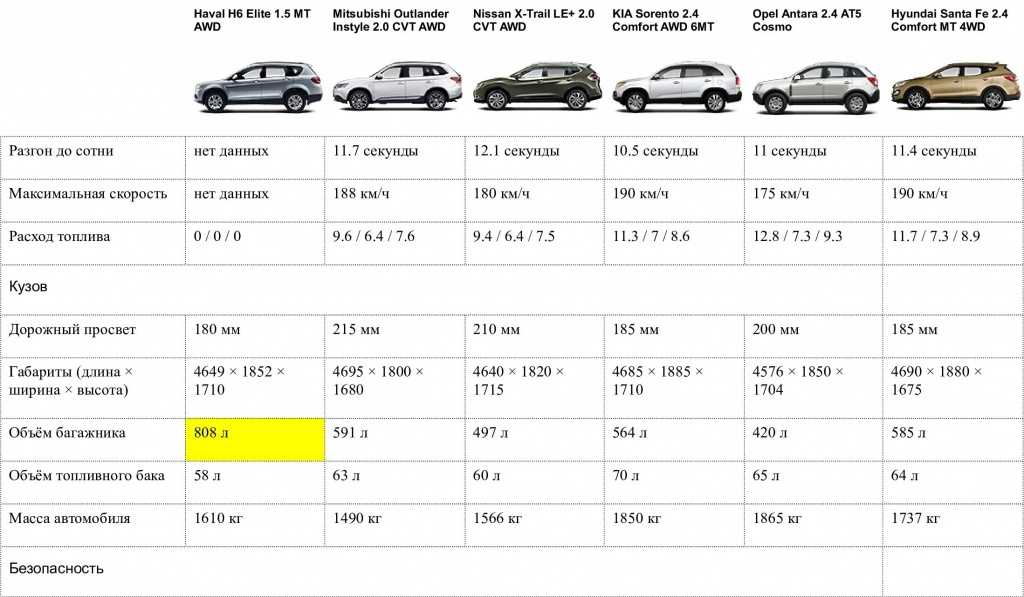
Так же, как люди группируют такие вещи, как мебель, фрукты или студенты колледжа. Вместе Кастак обучил морских львов мысленно сортировать символы по отдельным группы, закодированные цифрами и буквами. Для морских львов они просто черные метки на белых картах. Они должны были постепенно выучить ассоциации через годы проб и ошибок. Морские львы научились связывать группу из десяти букв вместе и группы из десяти цифр вместе, образуя связи между отдельными символами в каждой категории.
Рио преуспела в отслеживании этих групп. Для этого эксперимента она
вернулась к деревянной ширме у бассейна, где ей дали выбор между
буква и цифра в двух боковых окнах (центральное окно осталось
закрыто). Если, например, исследователи решили начать с букв
как правильный выбор, Рио была вознаграждена, когда впервые выбрала букву.
В ответ на эту награду она ткнула носом в письмо в последующем.
испытания. Заработав десять рыбок за десять ходов, правильный ответ поменялся местами.
к числам. Не зная об обратном, Рио неправильно выбрал букву.
Но уже на следующем испытании она изменила свою стратегию и начала выбирать
цифры и снова зарабатывать рыбу. «Этого нельзя было достичь
путем простых проб и ошибок, — сказал Кастак. «Это действительно
эмерджентное знание: они используют старую информацию по-новому.
Ее работа опубликована в Журнале экспериментального анализа
Поведение.
Заработав десять рыбок за десять ходов, правильный ответ поменялся местами.
к числам. Не зная об обратном, Рио неправильно выбрал букву.
Но уже на следующем испытании она изменила свою стратегию и начала выбирать
цифры и снова зарабатывать рыбу. «Этого нельзя было достичь
путем простых проб и ошибок, — сказал Кастак. «Это действительно
эмерджентное знание: они используют старую информацию по-новому.
Ее работа опубликована в Журнале экспериментального анализа
Поведение.
Еще одно новое измерение экспериментов включало изменение типа рыба используется для поощрения правильных ответов. Правильно подобранная буква заработала Рио кусок селедки, а номера были вознаграждены другим видом рыбы под названием мойва. В результате конкретное вознаграждение усиливало категории в ее уме.
Недавно Rio был протестирован с этой процедурой реверсирования после периода
год без проверки. Удивительно, но она выступила без единого изъяна. Она
вышло идеально. Я даже не мог в это поверить. Я сказал, что она должна была
хотя бы одну ошибку, потому что никто в это не поверит,
— с ухмылкой сказал Шустерман.
Она
вышло идеально. Я даже не мог в это поверить. Я сказал, что она должна была
хотя бы одну ошибку, потому что никто в это не поверит,
— с ухмылкой сказал Шустерман.
Чтобы еще раз проверить владение Рио в категориях, Кастака и Шустермана
добавлены новые символы в существующие наборы цифр и букв. Например,
они научили Рио тому, что новая буква, такая как «К», идет с
старый член буквенной группы, такой как J. Через пробы и
ошибка с селедкой, чтобы усилить связь, два символа стали
эквивалент в сознании Рио. Они повторили ту же процедуру с новыми
числа. Следующим шагом было проверить, эквивалентны ли теперь новые символы.
членам соответствующего оригинального набора в уме Рио. К
Чтобы проверить это, Кастак вернулся к эксперименту с тремя окнами. С одним из
оригинальные буквы, такие как A, в центральном окне, Рио
был предоставлен выбор K или новый номер. Она выбрала письмо
немедленно и получил ее селедку.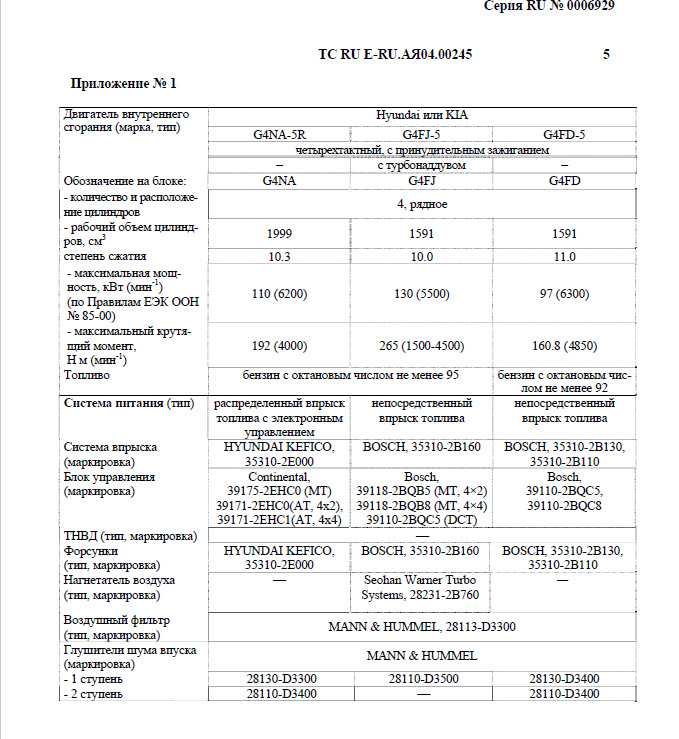 Не узнав напрямую, что K
был эквивалентен каждой из букв от A до J,
Рио смог провести корреляцию через одно соединение.
Не узнав напрямую, что K
был эквивалентен каждой из букв от A до J,
Рио смог провести корреляцию через одно соединение.
Результаты проливают свет на то, как способность логически мыслить может служить морской лев в природе. Это говорит о том, что животные могут делать выводы о вещах, которые мы никогда не понимали, они могли. Действительно логичные виды выводов, — сказал Шустерман.
Например, Рио смог связать различные сенсорные сигналы — вкус
взглядом друг на друга. В дикой природе эта способность может позволить
морского льва, чтобы мысленно сгруппировать вещи, такие как вид, запах и звук
друга или врага, что позволяет ей реагировать на любой из этих сигналов в
соответствующим образом. Например, когда мать возвращается домой после
двухдневное кормление, чтобы воссоединиться со своим щенком, они перекликаются
а затем использовать зрение и обоняние, чтобы подтвердить родство. Щенок может
узнает своих сестер таким же образом через их связи с
мать.
Морские львы также могут использовать этот навык для поиска пищи. Они могут связать еду с экологические подсказки, такие как образы и звуки, обычно связанные с еда. Эти подсказки могут привести их к еде в будущем. «Давайте» скажем, морской лев должен был добыть лосося с определенной лодки. Тогда это было бы свяжите этого лосося с лодкой и всеми аспектами лодки: конфигурация лодки, звуки лодки, время дня, когда кормление происходит эпизод. Итак, теперь лодка представляет собой лосося, — Шустерман. объяснил.
Другим важным результатом этого эксперимента было то, что он позволил
стареющий Рокки, чтобы присоединиться к Рио в ее способности замыкать логическую петлю. Пока
первая серия экспериментов часто вызывала разочарованный рев и метание
головы от Рокки, она преуспела в испытаниях с конкретной рыбой
награды. Ее выступление подкрепляет теории Шустермана о
практичность этих навыков. Это также снижает вероятность того, что
Предыдущий успех Рио был случайностью.
Это также снижает вероятность того, что
Предыдущий успех Рио был случайностью.
Хотя Рокки и Рио, несомненно, являются первопроходцами, они все еще только двое морские львы. Скептикам трудно оспаривать выводы Шустермана, но нужно больше работать с другими животными, прежде чем придет мысль, что животные способны к логике, могут широко применяться и восприниматься научное сообщество. Эту способность пытались выявить многие исследователи. у других животных. На сегодняшний день Шустерман и его морские львы одни в это достижение.
Морские львы умны. Но умнее ли они шимпанзе или дельфинов?
Более вероятным объяснением является метод, лежащий в основе «магии». В Центре умственной отсталости Шрайвера
работал с людьми, а также с обезьянами-капуцинами. Он считает, что Шустерман
тщательная методология может быть одним из ключей к его успеху с морскими львами.
«Рон думает как учитель, — сказал Макилвейн. У него есть
хорошее понимание того, что нужно сделать, чтобы результаты обучения были вероятными. Методы
требовательны к технике. Если бы лаборатории приматов использовали методы Рона,
они получат положительные результаты.
У него есть
хорошее понимание того, что нужно сделать, чтобы результаты обучения были вероятными. Методы
требовательны к технике. Если бы лаборатории приматов использовали методы Рона,
они получат положительные результаты.
Пока нет экспериментов, доказывающих, что приматы обладают способностью
мыслить логически, как Рокки и Рио, неофициальные данные свидетельствуют о том, что
Догадка Макилвейна верна. Херб Террас, психолог из Колумбийского университета
Университет сообщил об обучении шимпанзе языку жестов в 1986 году.
Шимпанзе научился подписывать слово «собака», когда увидел изображение собаки. затем
Однажды шимпанзе услышал собачий лай снаружи и подписал слово
собака.
Шустерман с оптимизмом смотрит на частичные успехи, наблюдаемые у таких видов, как
как приматы, дельфины и птицы. «Вполне вероятно, что животные
как у шимпанзе, если учесть тип обучения, который мы использовали, с
множество примеров сопоставления и обращения, было бы замечательно.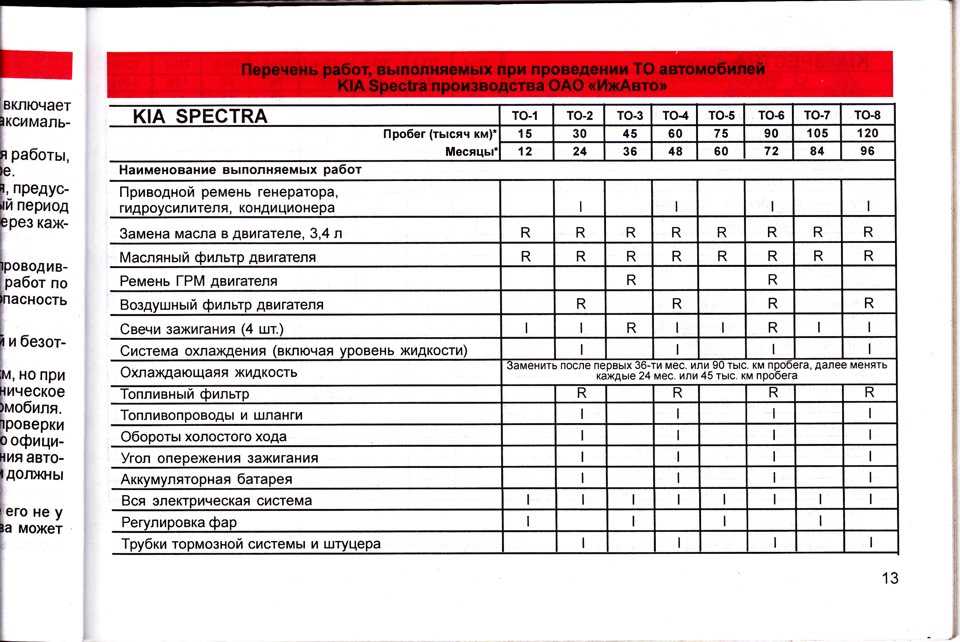 Успешная демонстрация логического мышления другими видами
поддерживает идею о том, что эквивалентность является эволюционным предшественником
язык.
Успешная демонстрация логического мышления другими видами
поддерживает идею о том, что эквивалентность является эволюционным предшественником
язык.
Следующим шагом Шустермана является использование того, что было изучено в лаборатории и применить его к наблюдениям в дикой природе. Ученые были очень сопротивляется идее, что морские львы являются социальными животными. Шустерман считает, что это может быть отчасти связано с трудностями наблюдения за морскими львами. в дикой природе.
Вместе с аспиранткой Эвелин Хангги Шустерман задокументировал дружеские
социальное поведение родственных самок калифорнийских морских львов в неволе
в парке морской жизни на острове Оаху. Научившись распознавать всех морских львов на
в парке они зафиксировали дружелюбное и агрессивное поведение. Они нашли
что почти все положительные взаимодействия, такие как плавание или дневной сон,
вместе, имели место между матерями и дочерьми, в то время как агрессивное поведение
был направлен на неродственных женщин.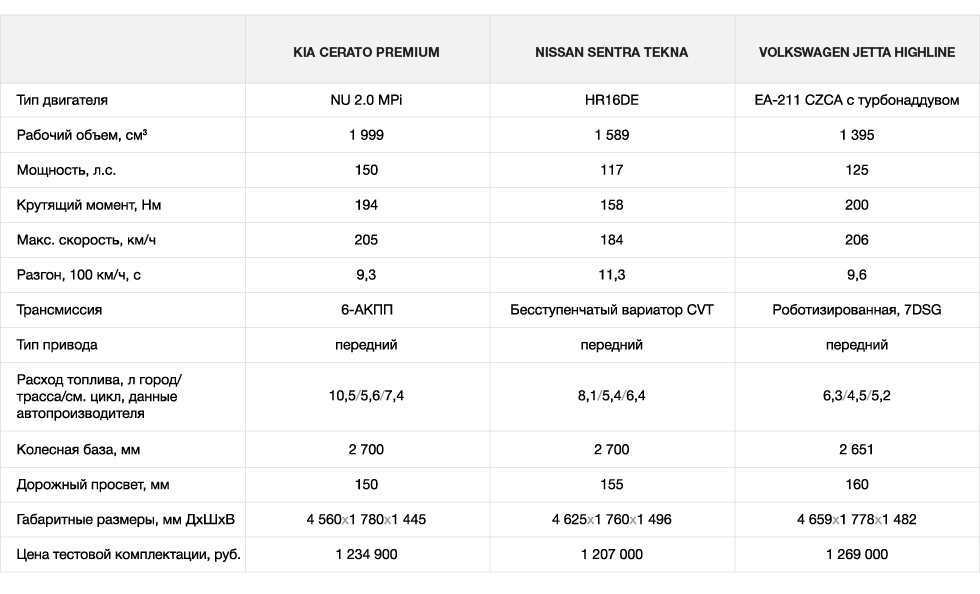 Но подобных работ еще предстоит
сделано в дикой природе. «ДНК — это ключ ко всему», — Шустерман.
сказал. Идея состоит в том, чтобы делать наблюдения и прогнозы относительно
родства, а затем проверить эти предсказания с помощью ДНК.
Но подобных работ еще предстоит
сделано в дикой природе. «ДНК — это ключ ко всему», — Шустерман.
сказал. Идея состоит в том, чтобы делать наблюдения и прогнозы относительно
родства, а затем проверить эти предсказания с помощью ДНК.
Шустерман положил глаз на особую социальную группу калифорнийских морских львов в Галапагосские острова, где он планирует проверить свои идеи. Естьs мало что известно о социальной организации морских львов, — сказал он. Еще нет.
Пищевые сигналы влияют на торможение моторных реакций и самооценку успеха диеты: графическое задание на аффективное переключение. Следовательно, постоянная саморегуляция автоматических склонностей к потреблению этих продуктов неизбежна для предотвращения увеличения веса (Lowe, 2003; Cohen and Farley, 2008). Важной предпосылкой успешной саморегуляции являются исполнительные функции (Hofmann et al., 2012). Недавние исследования были сосредоточены на трех основных аспектах управляющих функций: (1) рабочей памяти, (2) торможении и (3) когнитивной гибкости (Hofmann et al.
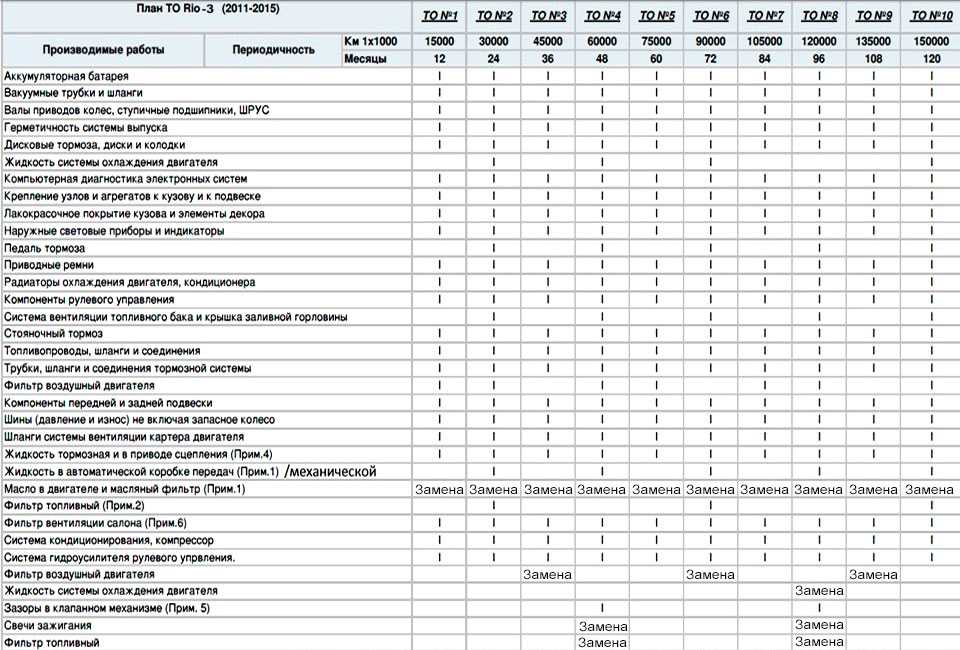 , 2012; Diamond, 2013). Рабочая память связана с сохранением и обновлением релевантной информации, сдерживанием сдерживания преобладающих импульсов, а когнитивная гибкость включает в себя изменение умственной установки, например, изменение точек зрения или подходов к проблеме и гибкое приспособление к новым требованиям, правилам или приоритетам (Hofmann et al. и др., 2012; Даймонд, 2013). Появляющиеся данные свидетельствуют о том, что эти исполнительные функции поддерживают важные механизмы в достижении целей саморегуляции и что временное снижение исполнительной функции может быть распространенным механизмом, способствующим нарушению саморегуляции (Hofmann et al., 2012).
, 2012; Diamond, 2013). Рабочая память связана с сохранением и обновлением релевантной информации, сдерживанием сдерживания преобладающих импульсов, а когнитивная гибкость включает в себя изменение умственной установки, например, изменение точек зрения или подходов к проблеме и гибкое приспособление к новым требованиям, правилам или приоритетам (Hofmann et al. и др., 2012; Даймонд, 2013). Появляющиеся данные свидетельствуют о том, что эти исполнительные функции поддерживают важные механизмы в достижении целей саморегуляции и что временное снижение исполнительной функции может быть распространенным механизмом, способствующим нарушению саморегуляции (Hofmann et al., 2012). В области пищевого поведения было показано, что пищевые сигналы и тяга к еде потребляют ресурсы саморегуляции и снижают производительность рабочей памяти (Kemps and Tiggemann, 2010; Meule et al., 2012e). Следовательно, более высокий объем рабочей памяти может способствовать успешной саморегуляции, связанной с приемом пищи (Hofmann et al. , 2012). Что касается когнитивной гибкости, то мало исследований касалось ее связи с саморегуляцией (Hofmann et al., 2012). Например, было высказано предположение, что гибкое диетическое поведение связано с лучшей саморегуляцией, связанной с питанием, по сравнению с жесткой диетой (Meule et al., 2011c; Westenhoefer et al., 2013). Недавно Дельгадо-Рико и соавт. (2012) обнаружили, что более высокий индекс массы тела (ИМТ) был связан с когнитивной ригидностью, измеренной с помощью задачи переключения реакции Струпа.
, 2012). Что касается когнитивной гибкости, то мало исследований касалось ее связи с саморегуляцией (Hofmann et al., 2012). Например, было высказано предположение, что гибкое диетическое поведение связано с лучшей саморегуляцией, связанной с питанием, по сравнению с жесткой диетой (Meule et al., 2011c; Westenhoefer et al., 2013). Недавно Дельгадо-Рико и соавт. (2012) обнаружили, что более высокий индекс массы тела (ИМТ) был связан с когнитивной ригидностью, измеренной с помощью задачи переключения реакции Струпа.
Поведенческое торможение и пищевое поведение
В отличие от рабочей памяти и когнитивной гибкости, существует множество исследований, изучающих связь между поведенческим торможением и пищевым регулированием. Двумя наиболее часто используемыми задачами для измерения поведенческого торможения являются задачи «идти/не идти» и парадигма стоп-сигнала. Задания «годен/не годен» включают в себя инструкцию реагировать на определенный стимул (например, нажатием кнопки), но подавлять эту реакцию на другой стимул. В задачах со стоп-сигналом сигнал запуска подается при каждой попытке, но в меньшинстве испытаний стоп-сигнал подается вскоре после появления сигнала начала, указывающий, что в этой попытке не следует нажимать кнопку. Задержка стоп-сигнала регулируется динамически, и время реакции на стоп-сигнал рассчитывается с более высокими значениями, указывающими на более низкую эффективность торможения (Logan et al., 19).97). В дальнейшем мы будем использовать термины поведенческое торможение, торможение реакции и ингибиторный контроль взаимозаменяемо в смысле отражения количества совершенных ошибок в задачах «годен/не годен» или времени реакции на стоп-сигнал в задачах на стоп-сигнал 1 . Кроме того, в задачах на поведенческое торможение могут быть рассчитаны другие показатели выполнения задачи, например, время реакции при правильных попытках и ошибках пропуска. Хотя ошибки пропуска, вероятно, отражают потерю внимания, интерпретация времени реакции не является однозначной, поскольку она не представляет собой ни чистую меру поведенческого торможения, ни отдельные механизмы внимания (Schulz et al.
В задачах со стоп-сигналом сигнал запуска подается при каждой попытке, но в меньшинстве испытаний стоп-сигнал подается вскоре после появления сигнала начала, указывающий, что в этой попытке не следует нажимать кнопку. Задержка стоп-сигнала регулируется динамически, и время реакции на стоп-сигнал рассчитывается с более высокими значениями, указывающими на более низкую эффективность торможения (Logan et al., 19).97). В дальнейшем мы будем использовать термины поведенческое торможение, торможение реакции и ингибиторный контроль взаимозаменяемо в смысле отражения количества совершенных ошибок в задачах «годен/не годен» или времени реакции на стоп-сигнал в задачах на стоп-сигнал 1 . Кроме того, в задачах на поведенческое торможение могут быть рассчитаны другие показатели выполнения задачи, например, время реакции при правильных попытках и ошибках пропуска. Хотя ошибки пропуска, вероятно, отражают потерю внимания, интерпретация времени реакции не является однозначной, поскольку она не представляет собой ни чистую меру поведенческого торможения, ни отдельные механизмы внимания (Schulz et al..jpg) , 2007).
, 2007).
Ряд исследований показал, что снижение способности к торможению связано с перееданием (см. обзор Guerrieri et al., 2008). Более низкая поведенческая заторможенность была продемонстрирована у пациентов с нервной булимией (Rosval et al., 2006; Wu et al., 2013), ограниченных в еде (Nederkoorn et al., 2004) и детей с ожирением (Nederkoorn et al., 2006a, 2007). и взрослые (Nederkoorn et al., 2006b) по сравнению с контрольной группой. Кроме того, было обнаружено, что торможение реакции приводит к умеренному потреблению пищи, так что в лабораторных условиях особенно те, кто ограничивал прием пищи и демонстрировал низкое торможение, ели больше (Jansen et al., 2009).; Меуле и др., 2011а). Следовательно, можно сделать вывод, что более низкое поведенческое торможение связано с более низкой саморегуляцией, связанной с приемом пищи, что проявляется в более высоком лабораторном потреблении пищи, более высоком ИМТ или переедании. Эти результаты, однако, не могут быть подтверждены рядом исследований, в которых не было обнаружено снижения выполнения заданий типа «давай/не ходи» или стоп-сигналов у пациентов с булимией, компульсивным перееданием или ожирением по сравнению с контрольной группой (Claes et al. , 2006, 2012; Galimberti et al., 2012; Hendrick et al., 2012; Van den Eynde et al., 2012; Wu et al., 2013).
, 2006, 2012; Galimberti et al., 2012; Hendrick et al., 2012; Van den Eynde et al., 2012; Wu et al., 2013).
В своей гедонистически-ингибирующей модели Appelhans (2009) интегрировал мотивацию аппетита в связи с едой (то, что он назвал гедонистическим кормлением ) и подавление этой мотивации аппетита. В соответствии с предположением, что эти два механизма взаимодействуют в контроле потребления пищи, Appelhans et al. (2011) смогли показать, что интерактивный эффект высокой чувствительности к пищевому вознаграждению (измеряемой с помощью шкалы Power of Food Scale ) и низкого ингибирующего контроля (измеряемой с помощью задачи дисконтирования с задержкой) предсказывает лабораторное потребление вкусных пищевых продуктов при избыточном весе и тучные женщины. Подобные интерактивные эффекты были обнаружены другими, например, низкий тормозной контроль (измеряемый с помощью парадигмы стоп-сигнала) в сочетании с высоким имплицитным предпочтением закусок предсказывал потребление лабораторных конфет (Hofmann et al.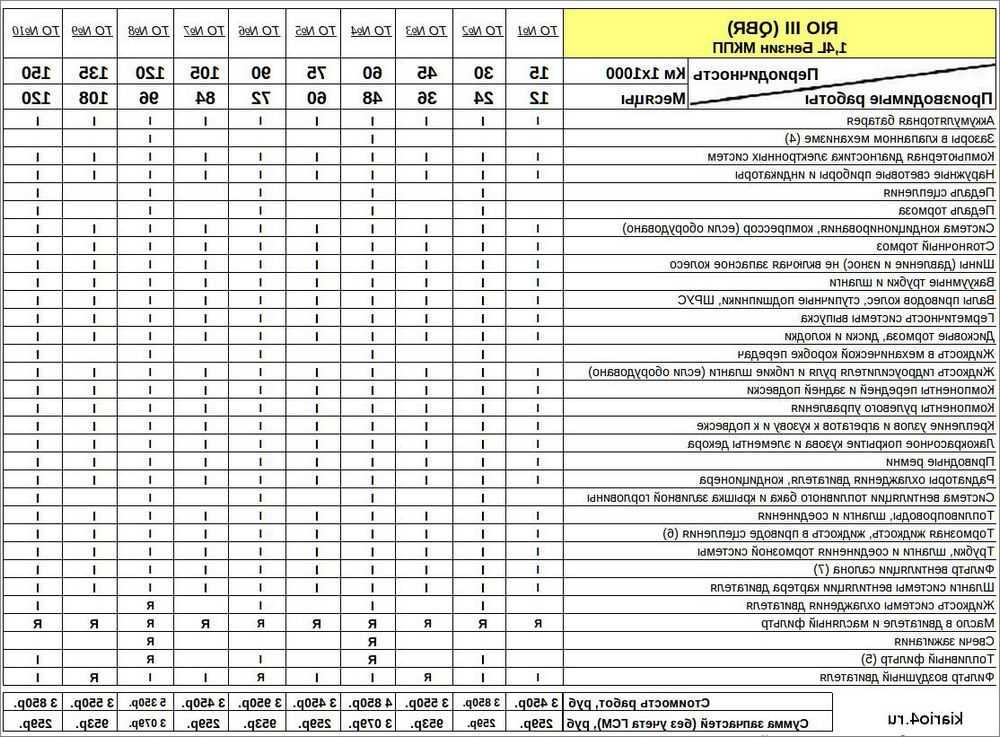 , 2009).) и увеличение веса в течение 1 года (Nederkoorn et al., 2010). Поскольку во всех этих исследованиях оценивалась общая способность к тормозному контролю, Аппельханс и его коллеги пришли к выводу, что «[…] явно необходимо разработать задачи, специально измеряющие тормозной контроль в контексте пищевого вознаграждения» (Appelhans et al. , 2011, стр. 2180). Это мнение было поддержано другими исследователями, например, в контексте исследования нейровизуализации в попытке «[…] разрушить механизмы, лежащие в основе чрезмерного потребления пищи, путем выхода за пределы парадигм приема пищи и пищевых сигналов в область когнитивных задач, предназначенных для выявлять импульсивное поведение, такое как задание «да/нет» […]» (Carnell et al., 2012, стр. 54).
, 2009).) и увеличение веса в течение 1 года (Nederkoorn et al., 2010). Поскольку во всех этих исследованиях оценивалась общая способность к тормозному контролю, Аппельханс и его коллеги пришли к выводу, что «[…] явно необходимо разработать задачи, специально измеряющие тормозной контроль в контексте пищевого вознаграждения» (Appelhans et al. , 2011, стр. 2180). Это мнение было поддержано другими исследователями, например, в контексте исследования нейровизуализации в попытке «[…] разрушить механизмы, лежащие в основе чрезмерного потребления пищи, путем выхода за пределы парадигм приема пищи и пищевых сигналов в область когнитивных задач, предназначенных для выявлять импульсивное поведение, такое как задание «да/нет» […]» (Carnell et al., 2012, стр. 54).
Поведенческое торможение и пищевые сигналы
В дополнение к тем исследованиям, которые связывают выполнение заданий на торможение двигательных реакций, не связанных с едой, с пищевым поведением, есть некоторые исследования, которые расширяют эти задачи с одновременным представлением пищевых сигналов (табл. 1). Таким образом, эти задачи измеряют реактивное торможение поведения, которое относится к прерыванию текущего поведения снизу вверх из-за условий мотивации, связанных с задачей (Nigg, 2000; Schulz et al., 2007; Claes et al., 2012). Согласно концепции Аппельханса (Appelhans, 2009 г.; Appelhans et al., 2011) можно было бы ожидать, что люди, склонные к перееданию, например, неудачно сидящие на диете или люди с ожирением или компульсивным перееданием, должны проявлять расторможенность именно в ответ на пищевые сигналы в задачах торможения моторных реакций из-за их низкий тормозной контроль и высокая чувствительность к пищевому вознаграждению. Однако исследования, в которых изучалось подавление поведения, вызванное пищевыми сигналами, довольно противоречивы. То есть некоторые исследования не выявили влияния пищевых сигналов на поведенческое торможение, некоторые обнаружили влияние на всех участников, а некоторые обнаружили влияние только на определенную подгруппу участников (таблица 1).
1). Таким образом, эти задачи измеряют реактивное торможение поведения, которое относится к прерыванию текущего поведения снизу вверх из-за условий мотивации, связанных с задачей (Nigg, 2000; Schulz et al., 2007; Claes et al., 2012). Согласно концепции Аппельханса (Appelhans, 2009 г.; Appelhans et al., 2011) можно было бы ожидать, что люди, склонные к перееданию, например, неудачно сидящие на диете или люди с ожирением или компульсивным перееданием, должны проявлять расторможенность именно в ответ на пищевые сигналы в задачах торможения моторных реакций из-за их низкий тормозной контроль и высокая чувствительность к пищевому вознаграждению. Однако исследования, в которых изучалось подавление поведения, вызванное пищевыми сигналами, довольно противоречивы. То есть некоторые исследования не выявили влияния пищевых сигналов на поведенческое торможение, некоторые обнаружили влияние на всех участников, а некоторые обнаружили влияние только на определенную подгруппу участников (таблица 1).
Таблица 1. Исследования, в которых изучалось влияние пищевых сигналов на поведенческое торможение .
Исследования, в которых изучалось влияние пищевых сигналов на поведенческое торможение .
Во-первых, следует отметить, что исследования существенно различаются по методологии, например, по типу исследуемых образцов (различаются по психопатологии, массе тела, возрасту или полу), типу используемого задания (годен/не годен по сравнению со стоп-сигналом) или дизайн исследования и представление стимулов (слова, изображения, экспозиция реальной пищи, положение стимулов на экране и т. д.). Во-вторых, в некоторых исследованиях показатели поведенческого торможения в ответ на пищевые сигналы не анализировались (или не сообщались), поскольку задание использовалось только в качестве фазы обучения или индукции торможения (Houben, 2011; Houben and Jansen, 2011; Veling et al. ., 2011). В-третьих, в одном исследовании была обнаружена положительная связь между количеством совершенных ошибок в ответ на изображения десертов и ИМТ (Batterink et al., 2010), а в другом исследовании количество совершенных ошибок было положительно связано с оценками эмоционального питания и отрицательно с выбором здоровой пищи (Jasinska et al.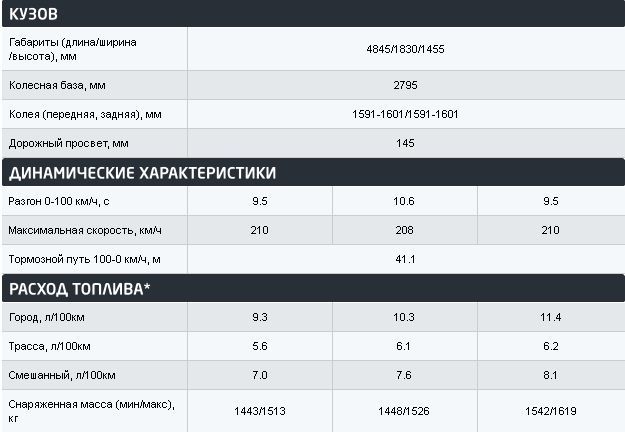 , 2012). Однако ни в одном из этих исследований не было контрольного условия, а это означает, что остается неясным, действительно ли на поведенческое торможение влияли пищевые стимулы или можно было бы получить такие же результаты с нейтральным заданием «да-нет». В-четвертых, в некоторых исследованиях было обнаружено влияние пищевых сигналов на время реакции или групповые различия в общем поведенческом торможении, но не было обнаружено специфического влияния пищевых сигналов (или воздействия пищи) на поведенческое торможение (Nederkoorn et al., 2004; Meule et al. , 2011а, 2012в). Наконец, в некоторых исследованиях можно было наблюдать влияние пищевых сигналов на торможение поведения (табл. 1). Используя нейтральную и специфичную для пищи задачу на стоп-сигнал, Houben et al. (2012b) обнаружили, что люди, не соблюдающие диету, были менее подавлены поведением в ответ на пищевые сигналы после воздействия пищи по сравнению с контрольным состоянием, и этот эффект не мог быть обнаружен у успешных людей, сидящих на диете, или в задаче с нейтральным стоп-сигналом.
, 2012). Однако ни в одном из этих исследований не было контрольного условия, а это означает, что остается неясным, действительно ли на поведенческое торможение влияли пищевые стимулы или можно было бы получить такие же результаты с нейтральным заданием «да-нет». В-четвертых, в некоторых исследованиях было обнаружено влияние пищевых сигналов на время реакции или групповые различия в общем поведенческом торможении, но не было обнаружено специфического влияния пищевых сигналов (или воздействия пищи) на поведенческое торможение (Nederkoorn et al., 2004; Meule et al. , 2011а, 2012в). Наконец, в некоторых исследованиях можно было наблюдать влияние пищевых сигналов на торможение поведения (табл. 1). Используя нейтральную и специфичную для пищи задачу на стоп-сигнал, Houben et al. (2012b) обнаружили, что люди, не соблюдающие диету, были менее подавлены поведением в ответ на пищевые сигналы после воздействия пищи по сравнению с контрольным состоянием, и этот эффект не мог быть обнаружен у успешных людей, сидящих на диете, или в задаче с нейтральным стоп-сигналом.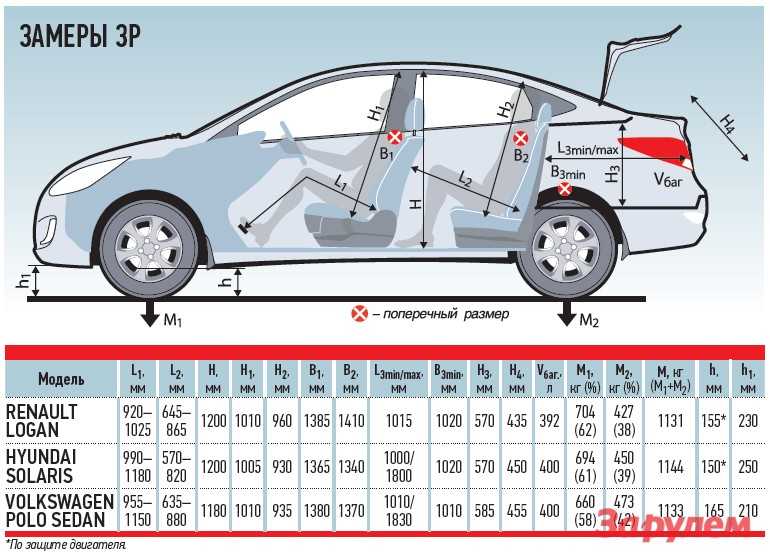 В другой задаче со стоп-сигналом Nederkoorn et al. (2012) обнаружили, что у детей с особенно избыточным весом наблюдалось меньшее торможение реакции на изображения еды по сравнению с изображениями игрушек. Совсем недавно более высокий SSRT в ответ на изображения еды был связан с более сильным желанием поесть после задания у молодых женщин (Meule et al., 2014).
В другой задаче со стоп-сигналом Nederkoorn et al. (2012) обнаружили, что у детей с особенно избыточным весом наблюдалось меньшее торможение реакции на изображения еды по сравнению с изображениями игрушек. Совсем недавно более высокий SSRT в ответ на изображения еды был связан с более сильным желанием поесть после задания у молодых женщин (Meule et al., 2014).
Задания на эмоциональное переключение, связанные с едой
В четырех исследованиях использовалось лексическое задание «да/нет», в котором участники должны были реагировать либо на еду, либо на нейтральные слова, но подавлять реакции на другую категорию. Картирование стимул-реакция переворачивается после каждого второго блока, и, следовательно, это задание измеряет как поведенческое торможение, так и когнитивную гибкость ( задание на аффективное переключение , AST; Mobbs et al., 2008, 2011). Снижение поведенческого торможения в этой задаче индексируется увеличением количества совершенных ошибок. Когнитивную гибкость можно оценить по общим различиям в выполнении задач между 9 детьми. 0887 сдвига и не сдвига блоков (см. описание AST ниже).
0887 сдвига и не сдвига блоков (см. описание AST ниже).
В первом исследовании все участники реагировали на пищу быстрее, чем на нейтральные мишени (Mobbs et al., 2008). Кроме того, у пациентов с нервной булимией наблюдалось более низкое торможение реакции (как при пищевом, так и при нейтральном блоках) по сравнению с контрольной группой, но не по АСТ, связанному с телом (Mobbs et al., 2008). Во втором исследовании снова все участники быстрее реагировали на пищевые мишени по сравнению с нейтральными мишенями (Mobbs et al., 2011). Участники с ожирением обычно совершали больше ошибок, чем контрольная группа (Mobbs et al., 2011). Важно отметить, что все участники сделали больше комиссионных ошибок в ответ на нейтральные слова (т. е. когда целевыми словами были слова о еде) по сравнению со словами о еде (т. е. когда нейтральные слова были целевыми) (Mobbs et al., 2011). В третьем исследовании было подтверждено, что все участники быстрее реагировали на слова о еде по сравнению со словами-объектами (Loeber et al.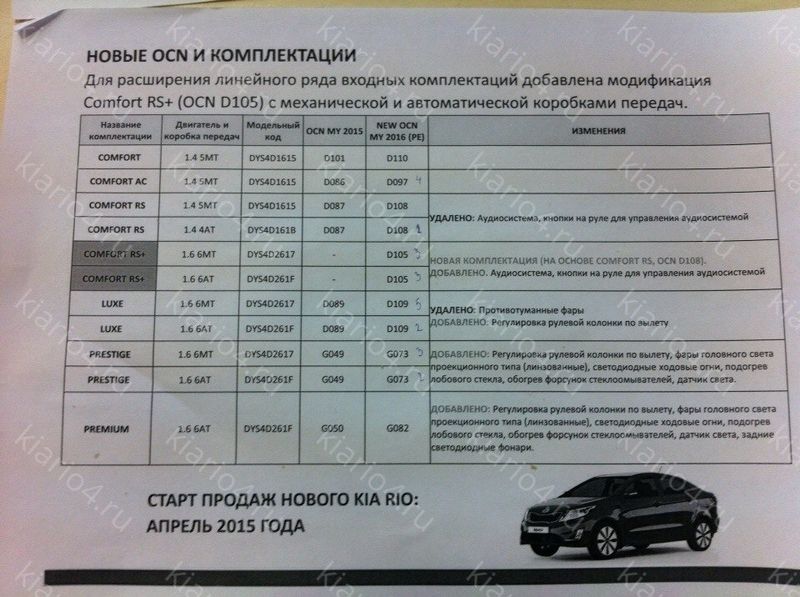 , 2012). Однако в этом исследовании участники допустили больше комиссионных ошибок в ответ на слова о еде (т. е. когда целью были нейтральные слова) по сравнению с нейтральными словами (т. е. когда мишенью были слова о еде) (Loeber et al., 2012). Различий между участниками с нормальным весом и ожирением обнаружено не было (Loeber et al., 2012). Совсем недавно у людей с нормальным весом можно было воспроизвести, что количество комиссионных ошибок было выше в ответ на пищу, чем на нейтральные слова (Loeber et al., 2013). Этот эффект был особенно выражен у участников с высокой субъективной оценкой голода, но не был связан с уровнем глюкозы в крови (Loeber et al., 2013).
, 2012). Однако в этом исследовании участники допустили больше комиссионных ошибок в ответ на слова о еде (т. е. когда целью были нейтральные слова) по сравнению с нейтральными словами (т. е. когда мишенью были слова о еде) (Loeber et al., 2012). Различий между участниками с нормальным весом и ожирением обнаружено не было (Loeber et al., 2012). Совсем недавно у людей с нормальным весом можно было воспроизвести, что количество комиссионных ошибок было выше в ответ на пищу, чем на нейтральные слова (Loeber et al., 2013). Этот эффект был особенно выражен у участников с высокой субъективной оценкой голода, но не был связан с уровнем глюкозы в крови (Loeber et al., 2013).
Подводя итог, можно сказать, что исследования с использованием АСТ, связанных с пищевыми продуктами, действительно выявили влияние пищевых сигналов на поведенческое торможение. Тем не менее, несколько вопросов остаются нерешенными. Во-первых, несмотря на то, что между едой и нейтральными мишенями были обнаружены различия в выполнении задач, вопреки ожиданиям, этот эффект не был связан с массой тела или особенностями пищевого поведения. В результате нельзя исключать альтернативные объяснения, такие как простой эффект размера категории (см. Обсуждение). Во-вторых, исследования противоречивы, поскольку неясно, может ли поведенческая расторможенность наблюдаться конкретно в ответ на слова о еде (хотя участников проинструктировали подавлять эти реакции) (Loeber et al., 2012, 2013) или в ответ на нейтральные отвлекающие факторы (когда участники должны реагировать на слова о еде) (Mobbs et al., 2011). В-третьих, неясно, как ингибирование реакции, связанной с пищей, связано с текущим статусом питания, поскольку оно было связано с самооценкой чувства голода, но не с уровнем глюкозы в крови (Loeber et al., 2013).
В результате нельзя исключать альтернативные объяснения, такие как простой эффект размера категории (см. Обсуждение). Во-вторых, исследования противоречивы, поскольку неясно, может ли поведенческая расторможенность наблюдаться конкретно в ответ на слова о еде (хотя участников проинструктировали подавлять эти реакции) (Loeber et al., 2012, 2013) или в ответ на нейтральные отвлекающие факторы (когда участники должны реагировать на слова о еде) (Mobbs et al., 2011). В-третьих, неясно, как ингибирование реакции, связанной с пищей, связано с текущим статусом питания, поскольку оно было связано с самооценкой чувства голода, но не с уровнем глюкозы в крови (Loeber et al., 2013).
В текущих исследованиях мы стремились прояснить и расширить эти предыдущие выводы. С этой целью мы использовали связанную с едой ТЧА, аналогичную описанным выше исследованиям (Mobbs et al., 2008, 2011; Loeber et al., 2012, 2013), но использовали изображения еды и нейтральных объектов, так как изображения еды были обнаружены оказывать более сильное воздействие по сравнению с пищевыми словами в когнитивных задачах (Brooks et al. , 2011). Основываясь на предыдущих выводах, мы ожидали, что участники будут быстрее реагировать на пищевые цели, чем на нейтральные цели. Кроме того, мы ожидали, что будут различия в поведенческом торможении между типами стимулов, которые могут проявляться в более высокой степени растормаживания (т. , 2012). Наконец, с помощью корреляционного и регрессионного анализов мы исследовали, связано ли выполнение задачи с голоданием, текущим пристрастием к еде, ИМТ, самооценкой успехов в соблюдении диеты и импульсивностью.
, 2011). Основываясь на предыдущих выводах, мы ожидали, что участники будут быстрее реагировать на пищевые цели, чем на нейтральные цели. Кроме того, мы ожидали, что будут различия в поведенческом торможении между типами стимулов, которые могут проявляться в более высокой степени растормаживания (т. , 2012). Наконец, с помощью корреляционного и регрессионного анализов мы исследовали, связано ли выполнение задачи с голоданием, текущим пристрастием к еде, ИМТ, самооценкой успехов в соблюдении диеты и импульсивностью.
Исследование 1
Материалы и методы
Участники
Участники женского пола были набраны среди студентов Вюрцбургского университета, Германия. Рекламные объявления были размещены в кампусе и с использованием списка рассылки студенческого совета. С женщинами, откликнувшимися на рекламу, связались по телефону ( N = 82) и проверили на предмет критериев исключения, включая психические расстройства, прием психотропных препаратов, недостаточный или избыточный вес (ИМТ < 17,5 или > 25 кг/м 9 ). 0760 2 ) и возраст > 40 лет. В исследование были включены только женщины с нормальным весом, потому что лишь немногие участницы скрининговой выборки имели избыточный вес, и, следовательно, распределение ИМТ было бы неравномерным. По той же причине подходящими считались только женщины моложе 40 лет. Всего в исследовании приняли участие n = 50 участников. Средний возраст составил M = 22,32 года ( SD = 3,03), а средний ИМТ M = 21,45 кг/м 2 ( SD = 2,67). Восемнадцать участников указали, что в настоящее время они пытаются контролировать свой вес (то есть сидят на диете). Пятеро участников были курильщиками. Пищевая депривация (т.е. среднее время с момента последнего приема пищи) составила M = 5,20 ч ( SD = 2,81). Участники либо получали кредиты за курс, либо 20 евро за участие 2 . . PSRS (Fishbach et al., 2003) использовался для оценки успеха диеты. В этой анкете из трех пунктов участники должны оценить по 7-балльной шкале, насколько успешно они следят за своим весом, худеют и насколько им сложно оставаться в форме.
0760 2 ) и возраст > 40 лет. В исследование были включены только женщины с нормальным весом, потому что лишь немногие участницы скрининговой выборки имели избыточный вес, и, следовательно, распределение ИМТ было бы неравномерным. По той же причине подходящими считались только женщины моложе 40 лет. Всего в исследовании приняли участие n = 50 участников. Средний возраст составил M = 22,32 года ( SD = 3,03), а средний ИМТ M = 21,45 кг/м 2 ( SD = 2,67). Восемнадцать участников указали, что в настоящее время они пытаются контролировать свой вес (то есть сидят на диете). Пятеро участников были курильщиками. Пищевая депривация (т.е. среднее время с момента последнего приема пищи) составила M = 5,20 ч ( SD = 2,81). Участники либо получали кредиты за курс, либо 20 евро за участие 2 . . PSRS (Fishbach et al., 2003) использовался для оценки успеха диеты. В этой анкете из трех пунктов участники должны оценить по 7-балльной шкале, насколько успешно они следят за своим весом, худеют и насколько им сложно оставаться в форме.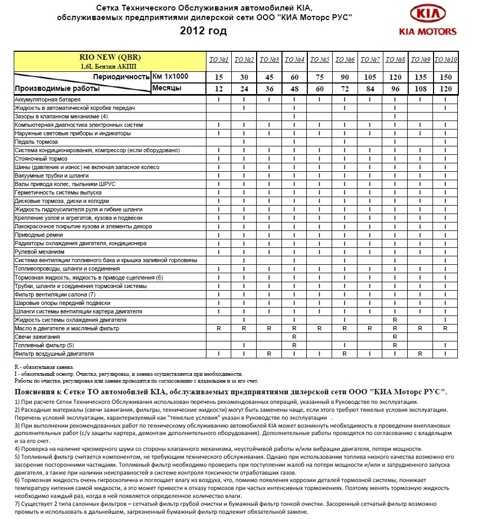 Валидность PSRS была продемонстрирована отрицательной связью с ИМТ, стратегиями жесткой диеты и другими показателями расторможенного питания, в то время как она положительно связана со стратегиями гибкой диеты (Meule et al., 2011c, 2012a, d). Внутренняя согласованность немецкой версии составляет α > 0,70 (Meule et al., 2012d) и составила α = 0,79.в текущем исследовании.
Валидность PSRS была продемонстрирована отрицательной связью с ИМТ, стратегиями жесткой диеты и другими показателями расторможенного питания, в то время как она положительно связана со стратегиями гибкой диеты (Meule et al., 2011c, 2012a, d). Внутренняя согласованность немецкой версии составляет α > 0,70 (Meule et al., 2012d) и составила α = 0,79.в текущем исследовании.
Шкала импульсивности Барратта — короткая версия (BIS-15) . BIS-15 был предложен Спинеллой (2007) в качестве укороченной версии BIS-11 (Patton et al., 1995) для измерения импульсивности по параметрам 90 887 моторной, 90 888 внимательной и 90 887 не 90 888 – 90 887 планирующей импульсивности 90 888. . Вместо 30 элементов, как в длинной версии, он состоит всего из 15 элементов. Конвергентная валидность BIS-15 была продемонстрирована связью от умеренной до сильной с Frontal Systems Behavior Scale и UPPS Impulsive Behavior Scale , в то время как дискриминантная валидность была отмечена слабой корреляцией с поиском ощущений (Spinella, 2007; Meule et al. , 2011b). Внутренняя согласованность немецкой версии α = 0,81 (Meule et al., 2011b). В текущем исследовании использовалась только общая оценка, а внутренняя согласованность составила α = 0,79.
, 2011b). Внутренняя согласованность немецкой версии α = 0,81 (Meule et al., 2011b). В текущем исследовании использовалась только общая оценка, а внутренняя согласованность составила α = 0,79.
Опросник пристрастия к еде — штат (FCQ-S) . FCQ-S (Cepeda-Benito et al., 2000) использовался для измерения текущей тяги к еде. Этот опросник из 15 пунктов оценивает мгновенную тягу к еде по параметрам: сильное желание поесть, ожидание положительного подкрепления, которое может возникнуть в результате еды, ожидание облегчения от негативных состояний и чувств в результате еды, отсутствие контроля над едой и тяга к пище. физиологическое состояние (Cepeda-Benito et al., 2000). На достоверность FCQ-S указывают положительные ассоциации с продолжительностью голодания и текущим негативным аффектом (Cepeda-Benito et al., 2003; Meule et al., 2012a). Кроме того, было обнаружено, что FCQ-S чувствителен к потреблению пищи и воздействию пищевых сигналов, так что тяга к состоянию снижается после завтрака (Cepeda-Benito et al. , 2000; Vander Wal et al., 2007) и увеличивается после выполнения упражнения. когнитивная задача с изображением еды (Meule et al., 2012e). Подшкалы сильно взаимосвязаны, и внутренняя согласованность общего балла составляет α = 0,9.2 (Меуле и др., 2012а). Поэтому мы использовали только общий балл для нашего анализа, а внутренняя согласованность в текущем исследовании составляла α = 0,90.
, 2000; Vander Wal et al., 2007) и увеличивается после выполнения упражнения. когнитивная задача с изображением еды (Meule et al., 2012e). Подшкалы сильно взаимосвязаны, и внутренняя согласованность общего балла составляет α = 0,9.2 (Меуле и др., 2012а). Поэтому мы использовали только общий балл для нашего анализа, а внутренняя согласованность в текущем исследовании составляла α = 0,90.
Аффективное задание на переключение (AST)
AST — это задание типа «годен/не годен», которое ранее использовалось с использованием эмоциональных (Murphy et al., 1999), связанных с алкоголем (Noël et al., 2005, 2007; Adams et al., 2013) и слова, связанные с едой или телом (Mobbs et al., 2008, 2011; Loeber et al., 2012, 2013). В текущем исследовании мы использовали модификацию этой задачи с изображением еды и нейтральных объектов. Продукты питания представляли собой изображения высококалорийных, вкусных сладких и соленых продуктов. Нейтральные картины были обычными предметами домашнего обихода. Все изображения были отредактированы, чтобы сделать их однородными по цвету фона (рис. 1). Программа была скомпилирована с использованием E-prime 2.0 (Psychology Software Tools Inc., Питтсбург, Пенсильвания) и отображена на 22-дюймовом ЖК-экране TFT. Участникам было предложено как можно быстрее нажимать кнопку ответа при предъявлении цели, но воздерживаться от ответов на отвлекающие факторы. Задача была разделена на 16 блоков по 18 попыток в каждом (всего = 288 попыток). В каждом блоке каждая картинка показывалась один раз, то есть половина картинок были мишенями, а половина — дистракторами. Картинки предъявлялись одна за другой в течение 500 мс в случайном порядке. Пустой экран был представлен во время межпробного интервала в течение 900 мс или участники получили обратную связь в случае ложной реакции или упущения. Перед каждым блоком в качестве целевой категории указывалась либо пища (F), либо объекты (O). Порядок блоков был либо FFOOFFOOFFOOFFOO, либо OOFFOOFFOOFFOOFF (уравновешенный по субъектам).
Все изображения были отредактированы, чтобы сделать их однородными по цвету фона (рис. 1). Программа была скомпилирована с использованием E-prime 2.0 (Psychology Software Tools Inc., Питтсбург, Пенсильвания) и отображена на 22-дюймовом ЖК-экране TFT. Участникам было предложено как можно быстрее нажимать кнопку ответа при предъявлении цели, но воздерживаться от ответов на отвлекающие факторы. Задача была разделена на 16 блоков по 18 попыток в каждом (всего = 288 попыток). В каждом блоке каждая картинка показывалась один раз, то есть половина картинок были мишенями, а половина — дистракторами. Картинки предъявлялись одна за другой в течение 500 мс в случайном порядке. Пустой экран был представлен во время межпробного интервала в течение 900 мс или участники получили обратную связь в случае ложной реакции или упущения. Перед каждым блоком в качестве целевой категории указывалась либо пища (F), либо объекты (O). Порядок блоков был либо FFOOFFOOFFOOFFOO, либо OOFFOOFFOOFFOOFF (уравновешенный по субъектам). Из-за такого расположения четыре блока каждой целевой категории представляли собой 90 887 сдвиговых 90 888 блоков, в которых участники должны были обратить вспять ассоциации стимул-реакция предыдущего блока, и четыре блока представляли собой 90 887 несменных 90 888 блоков, в которых ассоциации стимул-реакция были одинаковыми. как в предыдущем блоке. Чтобы гарантировать, что первый блок может быть проанализирован как сменный блок, перед тестовыми блоками был запущен тренировочный блок с использованием противоположной целевой категории. Вся работа длилась около 10 мин.
Из-за такого расположения четыре блока каждой целевой категории представляли собой 90 887 сдвиговых 90 888 блоков, в которых участники должны были обратить вспять ассоциации стимул-реакция предыдущего блока, и четыре блока представляли собой 90 887 несменных 90 888 блоков, в которых ассоциации стимул-реакция были одинаковыми. как в предыдущем блоке. Чтобы гарантировать, что первый блок может быть проанализирован как сменный блок, перед тестовыми блоками был запущен тренировочный блок с использованием противоположной целевой категории. Вся работа длилась около 10 мин.
Рисунок 1. Стимулирующий набор из (A) изображений еды и (B) нейтральных изображений в исследовании 1 .
Процедура
Участники проходили тестирование с 9:00 до 18:00. (среднее время тестирования 12:00). Всех участников просили не употреблять пищу, кофеин, никотин или алкоголь как минимум за 3 часа до эксперимента. После того, как участники выполнили AST, они немедленно заполнили FCQ-S и сообщили о часах, прошедших с момента их последнего приема пищи.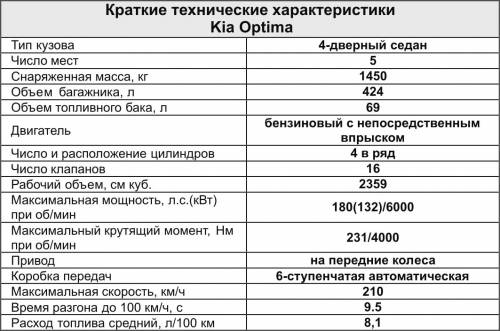 Заполнение остальных анкет и измерение роста и веса участников проводилось либо в тот же день, либо в течение 1–2 недель после эксперимента в зависимости от индивидуальной отнесенности к условиям эксперимента.
Заполнение остальных анкет и измерение роста и веса участников проводилось либо в тот же день, либо в течение 1–2 недель после эксперимента в зависимости от индивидуальной отнесенности к условиям эксперимента.
Анализ данных
Испытания со временем реакции менее 150 мс, отражающие ожидание, исключались из анализа. Показатели, представляющие интерес, представляли собой время реакции (мс) в пробных попытках (т. е. время, необходимое для ответа на каждую цель), количество совершенных ошибок (т. е. ответов на отвлекающие факторы) и ошибки упущения (т. е. неспособность реагировать на цели). Корреляции между различными показателями выполнения задач показаны в таблице 2.
Таблица 2. Коэффициенты корреляции Пирсона между показателями выполнения задания .
A 2 ( тип мишени : еда по сравнению с объектом) × 2 ( тип блока : сдвиг по сравнению с отсутствием смены) Дисперсионный анализ для повторных измерений был рассчитан для каждой зависимой переменной (время реакции, упущение и ошибки комиссии) .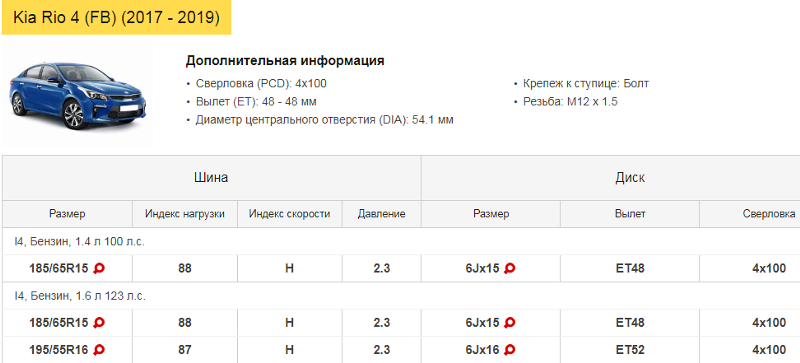 Post-hoc t — тесты рассчитывались в случае значительных взаимодействий. Кроме того, мы рассчитали корреляции между параметрами выполнения заданий в пищевых блоках, нейтральных блоках и баллами различий (еда минус нейтральные) с ИМТ, пищевой депривацией и показателями анкеты. Результаты считались значимыми при уровне α 9.0887 р = 0,05. Результаты, отмеченные как нс , относятся к p — значения > 0,05.
Post-hoc t — тесты рассчитывались в случае значительных взаимодействий. Кроме того, мы рассчитали корреляции между параметрами выполнения заданий в пищевых блоках, нейтральных блоках и баллами различий (еда минус нейтральные) с ИМТ, пищевой депривацией и показателями анкеты. Результаты считались значимыми при уровне α 9.0887 р = 0,05. Результаты, отмеченные как нс , относятся к p — значения > 0,05.
Результаты
Производительность задачи
Время реакции
Был значительный основной эффект для типа цели [ F (1, 49) = 202.11, P <0,001,
6.103487. = 0,81], что указывает на более быстрое время реакции на пищевые мишени ( M = 366,15 мс, SD = 16,60), чем на нейтральные мишени ( M = 387,98 мс, SD = 14,70). Ни главный эффект для блочного типа [ F (1, 49) = 0,05, нс , η 2 p = 0,00], ни тип взаимодействия целевого типа [
F 8 (1, 49) = 0,81, нс , η 2 p = 0,00] были значимыми (рис. 2А).
2А).Рис. 2. Выполнение задачи в зависимости от типа цели и типа блока в исследовании 1 . Отображается (A) время реакции в го-триалах в мс, (B) количество совершенных ошибок и (C) ошибок пропусков. Столбики погрешностей указывают стандартные ошибки среднего. *** р < 0,001, * р < 0,05.
Ошибки комиссий
Основной эффект для типа мишени был несущественным [ F (1, 49) = 0,54, ns , η 2
p8], но было а =
8
8 значительный главный эффект для блочного типа [ F (1, 49) = 23,82, P <0,001, η 2 P = 0,33] Указывание о большем количестве ошибок комиссии в сдвиге ( M = 0,33], указывая на большее количество ошибок комиссии в сменных блоках ( M = 0,33], указывая на большее количество ошибок комиссии в сдвиге ( M
, 0,33]. ), чем в блоках без сдвига ( M = 2,03, SD = 1,22). Тип мишени взаимодействия × тип блока не был значительным [ F (1, 49) = 3,84, нс , η 2 p = 0,07, рис. 2B].
Тип мишени взаимодействия × тип блока не был значительным [ F (1, 49) = 3,84, нс , η 2 p = 0,07, рис. 2B].Ошибки пропуска
Был значительный основной эффект для типа цели [ F (1, 49) = 56,65, P <0,001, η 2 P = 0,544] и блокирует тип = 0,544] и блокирует тип = 0,544] и блокирует тип P = 0,544] и блокирует тип P
= 0,544]. (1, 49) = 33,65, p < 0,001, η 2 p = 0,41], что указывает на большее количество ошибок пропуска в ответ на нейтральные цели ( M = 4,18, 9087), SD 9088 к пищевым мишеням ( M = 2,1, SD = 1,79) и больше ошибок пропуска в блоках сдвига ( M = 3,76, SD = 2,11), чем в блоках без сдвига ( M = 2,44, SD = 1,73). Эти основные эффекты были дополнительно квалифицированы по значительному взаимодействию тип мишени × тип блока [ F (1, 49) = 4,21, p < 0,05, η 2 p = 0,08]. Апостериорный -тест показал, что разница между блоками сдвига и блоками без сдвига была особенно заметна для нейтральных целей [ M (сменно-несменно) = 1,84, SD = 3,01] по сравнению с пищевыми целями [ M (сменно-несменно) = 0,80, SD = , t (49) = 2,05, р < 0,05, рис. 2С].
Апостериорный -тест показал, что разница между блоками сдвига и блоками без сдвига была особенно заметна для нейтральных целей [ M (сменно-несменно) = 1,84, SD = 3,01] по сравнению с пищевыми целями [ M (сменно-несменно) = 0,80, SD = , t (49) = 2,05, р < 0,05, рис. 2С].
Корреляционный анализ
ИМТ отрицательно коррелировал с успехом диеты ( r = -0,67, p < 0,001). ИМТ был положительным, а успех диеты отрицательно коррелировал с разницей в баллах за ошибки комиссии (таблица 3). При более высоком ИМТ и более низком успехе диеты участники совершали больше ошибок в пищевых блоках по сравнению с нейтральными блоками. Выполнение задания не было связано с пищевой депривацией, текущим пристрастием к еде и импульсивностью (таблица 3).
Таблица 3. Коэффициенты корреляции Пирсона между выполнением задания и ИМТ, пищевой депривацией и показателями анкеты (исследование 1) .
Заключение исследования 1
Исследование 1 показало, что участники быстрее реагировали на пищу, чем нейтральные мишени. Они также пропускали меньше еды, чем нейтральные цели. Ошибки комиссии не различались между пищевыми и нейтральными блоками. Таким образом, эти результаты показывают, что, хотя могут быть обнаружены различия в выполнении задач между пищевыми и нейтральными сигналами, пищевые сигналы не влияют на поведенческое торможение. Однако в целом участники допустили лишь несколько ошибок ( М = 5,44 ошибки в пищевых блоках и М = 5,02 ошибки в нейтральных блоках) и, таким образом, отсутствие разницы в поведенческом торможении может быть связано с эффектом потолка.
Более высокий ИМТ и более низкий успех диеты были связаны с более высокой расторможенностью (т.е. большим количеством ошибок) в еде по сравнению с нейтральными блоками. Пищевая депривация, тяга к еде после выполнения задания и импульсивность не были связаны с выполнением задания. Таким образом, довольно неудачный контроль над потреблением пищи, по-видимому, связан с нарушением поведенческого торможения при столкновении с вкусными пищевыми сигналами, и эта связь, по-видимому, не зависит от текущей депривации пищи, тяги к еде или импульсивности.
Таким образом, довольно неудачный контроль над потреблением пищи, по-видимому, связан с нарушением поведенческого торможения при столкновении с вкусными пищевыми сигналами, и эта связь, по-видимому, не зависит от текущей депривации пищи, тяги к еде или импульсивности.
Исследование 2
В исследовании 2 использовались те же показатели состояния и признаков [т. е. лишение пищи (время, прошедшее с момента последнего приема пищи), состояние тяги к еде, ИМТ, успешность диеты и импульсивность] и, опять же, наглядная версия был введен АСТ; тем не менее, в план исследования были внесены следующие изменения:
(1) Поскольку участники допустили лишь несколько ошибок в исследовании 1, количество испытаний было увеличено с 288 до 320, чтобы получить большее количество комиссионных ошибок.
(2) В исследовании 1, состояние тяги к еде после задание не было связано ни с выполнением задания, ни с ИМТ, ни с какими-либо показателями самоотчета. Тем не менее, состояние тяги к еде обычно увеличивается во время таких заданий (см. Meule et al., 2012e; Meule and Kübler, в редакции), и, следовательно, возможно, что тяга к до или увеличение во время задания может быть связано с выполнение задачи, ИМТ или показатели самоотчета. Так, в исследовании 2 состояние пищевого влечения оценивалось также до задания в дополнение к его оценке после задания.
Meule et al., 2012e; Meule and Kübler, в редакции), и, следовательно, возможно, что тяга к до или увеличение во время задания может быть связано с выполнение задачи, ИМТ или показатели самоотчета. Так, в исследовании 2 состояние пищевого влечения оценивалось также до задания в дополнение к его оценке после задания.
(3) Различия в выполнении задания, т. е. времени реакции и ошибках пропусков, между пищевыми и нейтральными стимулами в исследовании 1 могут быть обусловлены рядом причин, например, характеристиками стимулов (например, визуальная сложность, размер категории) или голод (например, участники быстрее реагируют на пищевые стимулы, когда они голодны). Таким образом, в исследовании 2 еда и нейтральные изображения были сопоставлены в отношении физических характеристик, таких как визуальная сложность. Кроме того, пищевым статусом манипулировали таким образом, что половина участников потребляла пищу в лаборатории (группа сытости, n = 51), а другая половина была лишена пищи в течение нескольких часов (голодная группа, n = 51).
Были сформулированы следующие гипотезы:
(1) Текущая тяга к еде : Потребление пищи уменьшит тягу к еде, о которой сообщают сами участники сытой группы, и, таким образом, сытая группа будет сообщать о более низкой тяге к еде, о чем свидетельствуют баллы на шкале. FCQ-S до и после задания по сравнению с голодной группой. Тем не менее, самооценка тяги к еде должна была быть выше после выполнения задания по сравнению с тем, что было до нее, в обеих группах, поскольку это обычно можно обнаружить при исследовании когнитивных задач с визуальными, вкусными пищевыми стимулами (Meule et al., 2012e; Meule and Kübler). , в редакции).
(2) Корреляты текущей тяги к еде : Хотя мы ожидали общего увеличения тяги к еде во время AST из-за воздействия пищевых сигналов, существуют также межиндивидуальные различия в реактивности пищевых сигналов. Например, более высокая реактивность пищевых сигналов наблюдалась у людей с более высоким ИМТ (Jansen et al., 2003), ограниченным приемом пищи (Fedoroff et al. , 1997) и более высокой импульсивностью (Tetley et al., 2010). . Кроме того, реакция на пищевые сигналы у довольно неудачно сидящих на диете увеличивается после приема высококалорийной предварительной нагрузки (Stroebe, 2008). Таким образом, мы исследовали, были ли изменения в состоянии тяги к еде во время ТЧА связаны с текущей депривацией пищи или переменными чертами (ИМТ, успешность диеты, импульсивность) и были ли эти ассоциации модулированы приемом пищи.
, 1997) и более высокой импульсивностью (Tetley et al., 2010). . Кроме того, реакция на пищевые сигналы у довольно неудачно сидящих на диете увеличивается после приема высококалорийной предварительной нагрузки (Stroebe, 2008). Таким образом, мы исследовали, были ли изменения в состоянии тяги к еде во время ТЧА связаны с текущей депривацией пищи или переменными чертами (ИМТ, успешность диеты, импульсивность) и были ли эти ассоциации модулированы приемом пищи.
(3) Выполнение задания : Мы ожидали повторить результаты исследования 1, согласно которым участники будут реагировать быстрее и пропускать меньше пищи, чем нейтральные мишени. По мере того, как мы увеличивали количество испытаний, мы ожидали, что участники будут совершать больше ошибок в целом, и, следовательно, теперь мы также обнаружим различия в ошибках комиссии между пищевыми и нейтральными блоками. Как отмечалось выше, существующая литература дала противоположные результаты, поэтому мы сформулировали ненаправленную гипотезу о том, что участники могут совершать больше ошибок в пищевых блоках (Mobbs et al. , 2011) или в нейтральных блоках (Loeber et al., 2012). Более того, недавно было обнаружено, что различия в ошибках совершения и пропусков между блоками с лексической пищей и нейтральными стимулами особенно заметны у людей с высоким самоотчетом о голоде (Loeber et al., 2013). Таким образом, мы ожидали, что в сытой группе различия в выполнении задач между пищевыми и нейтральными блоками будут ослаблены.
, 2011) или в нейтральных блоках (Loeber et al., 2012). Более того, недавно было обнаружено, что различия в ошибках совершения и пропусков между блоками с лексической пищей и нейтральными стимулами особенно заметны у людей с высоким самоотчетом о голоде (Loeber et al., 2013). Таким образом, мы ожидали, что в сытой группе различия в выполнении задач между пищевыми и нейтральными блоками будут ослаблены.
(4) Корреляты выполнения задачи : Основываясь на результатах исследования 1, мы ожидали, что более высокий ИМТ и более низкий успех диеты будут связаны с большим количеством ошибок в еде по сравнению с нейтральными блоками. Поскольку ожидалось, что участники в целом совершат больше ошибок, мы теперь предположили, что количество совершенных ошибок будет положительно связано с самооценкой импульсивности (Aichert et al., 2012). Все анализы проводились с использованием ошибок совершения, а также ошибок времени реакции и пропусков, чтобы выяснить, связаны ли результаты конкретно с поведенческим торможением или с общим выполнением задачи. Наконец, мы исследовали, будет ли выполнение задачи связано с переменными состояния, то есть с голодом и насыщением в настоящее время, и, возможно, связи между переменными состояния и черты с выполнением задачи будут модулироваться группой (то есть голодом и насыщением).
Наконец, мы исследовали, будет ли выполнение задачи связано с переменными состояния, то есть с голодом и насыщением в настоящее время, и, возможно, связи между переменными состояния и черты с выполнением задачи будут модулироваться группой (то есть голодом и насыщением).
Материалы и методы
Участники
Участники женского пола были набраны среди студентов Вюрцбургского университета, Германия. Всего в исследовании приняли участие N = 102 участника. Как и в исследовании 1, все участники были моложе 40 лет ( M = 22,76 года, SD = 3,72). Однако никаких ограничений в отношении ИМТ не вводилось. В результате средний ИМТ был немного выше, но все же сравним с исследованием 1 ( M = 22,11 кг/м 9 ).0760 2
, SD = 3,36). Тридцать один участник (30,4%) указали, что в настоящее время они пытаются контролировать свой вес (то есть сидят на диете). Восемь участников (7,8%) были курильщиками. Пищевая депривация (т.е. среднее время с момента последнего приема пищи) составила M = 7,5 ч ( SD = 4,93). Участники либо получали кредиты за курс, либо 6 евро за участие.
Участники либо получали кредиты за курс, либо 6 евро за участие.Анкеты
Использовались те же анкеты, что и в исследовании 1.
AST
Снова использовалась графическая версия задачи AST. Из 9 вариантов были выбраны 10 изображений продуктов питания и 10 нейтральных изображений (рис. 3).0887 база данных food.pics , которая включает, среди прочего, информацию о физических характеристиках (см. www.food-pics.sbg.ac.at; Meule and Blechert, 2012) 3 . Пищевые и нейтральные стимулы не различались по визуальной сложности (сжатие jpg, определение границ, субъективные оценки) и яркости и контрастности RGB (все t s (18) < 1,72, ns ). Задача была аналогична исследованию 1, за исключением того, что 16 блоков теперь состояли из 20 испытаний каждый (всего = 320 испытаний), а интервал между испытаниями был изменен на 1000 мс.
Рисунок 3. Стимулирующий набор из (A) изображений еды и (B) нейтральных изображений в исследовании 2 .
Процедура
Участники проходили тестирование с 8:00 до 20:00. (среднее время тестирования 12:45). Всех участников просили не принимать пищу как минимум за 3 часа до эксперимента. После того, как участники подписали информированное согласие, половина участников заполнили анкету FCQ-S и впоследствии выпили миску сливочного кефира (Desira ® калорий и питательных веществ на порцию (250 г): 360 ккал, 5,8 г белка, 42,3 г углеводов). , 18,8 г жира), вкус которого сравним со сладким фруктовым йогуртом. Перед AST все участники заполнили FCQ-S (т. е. половина участников сделала это в первый раз, а половина участников — во второй раз). После того, как участники выполнили AST, они сразу же снова заполнили FCQ-S и другие анкеты. Наконец, измеряли рост и вес участников.
Анализ данных
Текущая тяга к еде . Группы (т. е. голодные и сытые) сравнивали по возрасту, ИМТ и голоданию с помощью независимых t -тестов, а также по статусу соблюдения диеты и курения с помощью χ 2 -тестов. Исходные уровни тяги к еде (т. е. баллы FCQ-S непосредственно перед ТЧА в голодной группе и баллы FCQ-S перед приемом пищи в сытой группе) сравнивали с независимым тестом t . В сытой группе изменения показателей FCQ-S до и после приема пищи тестировались с помощью парного теста 9.0887 т -тест. Групповые различия и изменения до и после ТЧА были протестированы с помощью дисперсионного анализа 2 (между субъектами фактор группа : голодный и сытый) × 2 (внутрисубъектный фактор время : до и после задания) для повторных измерений .
Исходные уровни тяги к еде (т. е. баллы FCQ-S непосредственно перед ТЧА в голодной группе и баллы FCQ-S перед приемом пищи в сытой группе) сравнивали с независимым тестом t . В сытой группе изменения показателей FCQ-S до и после приема пищи тестировались с помощью парного теста 9.0887 т -тест. Групповые различия и изменения до и после ТЧА были протестированы с помощью дисперсионного анализа 2 (между субъектами фактор группа : голодный и сытый) × 2 (внутрисубъектный фактор время : до и после задания) для повторных измерений .
Корреляты текущей тяги к еде . Были исследованы взаимосвязи пищевой депривации, ИМТ, успешности диеты и импульсивности с состоянием тяги к еде (FCQ-S до задания, FCQ-S после задания и разница FCQ-S после задания минус FCQ-S до задания). с линейным регрессионным анализом. Для каждой из этих переменных была рассчитана регрессионная модель, включающая соответствующую переменную, а также группу и коэффициент взаимодействия обоих в качестве предикторов состояния тяги к еде.
Выполнение задачи . Расчет выполнения задания был аналогичен исследованию 1, и корреляции между индексами выполнения задания показаны в таблице 2. A 2 (внутрисубъектный фактор , тип цели : еда и объект) × 2 (внутрисубъектный фактор , тип блока : смена по сравнению с отсутствием смены) × 2 (между факторами субъекта и группой : голодные и сытые) ANOVA для повторных измерений рассчитывался для каждой зависимой переменной (время реакции, упущение и ошибки совершения). Post-hoc t — тесты рассчитывались в случае значительных взаимодействий.
Корреляты выполнения задания . Взаимосвязь пищевой депривации, состояния тяги к еде (т. е. разница баллов FCQ-S после задания минус баллы FCQ-S до задания), ИМТ, успешности диеты и импульсивности с выполнением задания изучали с помощью линейного регрессионного анализа. Для каждой из этих переменных была рассчитана регрессионная модель, включающая соответствующую переменную, а также группу (т. е. голодных и сытых) и условия взаимодействия обоих в качестве предикторов выполнения задачи.
е. голодных и сытых) и условия взаимодействия обоих в качестве предикторов выполнения задачи.
Результаты
Текущая тяга к еде
Группы не различались по возрасту, ИМТ, пищевой депривации [все t s (100) < 1,09, ns ] или пропорции тех, кто сидит на диете, и тех, кто курит χ 2 с (1) < 0,55, нс ]. Они также не различались по исходным уровням тяги, то есть когда FCQ-S был завершен в первый раз после прибытия в лабораторию [т. задание в голодной группе; t (100) = 0,65, нс ]. В сытой группе тяга уменьшалась после приема пищи (90 887 M 90 888 = 29,28, 90 887 SD 90 888 = 10,11) по сравнению с до [90 887 M 90 888 = 40,22, 90 887 SD 90 888 = 9,16, 90 887 t 90 888 90 340 (50 887 t 90 888 90 340). 10,19, р < 0,001]. ANOVA для тяги до и после задания выявил значимые основные эффекты для группы [ F (1, 100) = 37,31, p < 0,001, η 2 P = 0,27] и время [ F (1, 100) = 65,87, P <0,001, η 2 P 89960 2 P 89960 2 P 888910 2 P 8888910 2 P888888910. СО) по сравнению с предыдущим ( M = 35,36, SD = 11,80, рис. 4). Взаимодействие группа × время не значимо [ F (1, 100) = 0,67, нс ] 4 . Рисунок 4. Баллы по опроснику о пристрастии к еде — состояние в зависимости от группы до и после задания на аффективное переключение в исследовании 2 . Аналогично выводам, приведенным выше, группа предсказала тягу к еде до и после задания таким образом, что голодная группа сообщила о более высокой тяге к еде (Таблица 4). ИМТ негативно предсказывал тягу к еде до и после задания. Наконец, взаимодействие 9Группа 0887 × успех диеты достоверно предсказывал разницу между тягой к еде до и после задания: успех диеты отрицательно предсказывал усиление тяги к еде в группе, насытившейся (β = -0,32, p < 0,05), но не в голодная группа (β = 0,16, нс , рис. Таблица 4. Прогноз состояния тяги к еде в зависимости от пищевой депривации, ИМТ, успешности диеты, импульсивности и группы (исследование 2) . Рисунок 5. Состояние тяги к еде (баллы по Опроснику тяги к еде — состояние после задания минус баллы перед заданием) в зависимости от успеха группы и диеты в исследовании 2 . Все переменные z -стандартизированы. Был основной эффект для типа мишени [ F (1, 100) = 116,15, P <0,001, η 2 9 888 <0,001, η 2 9 888 <0,001, η 2 9 888. что указывает на то, что участники реагировали быстрее в ответ на пищевые цели ( M = 373,72 мс, SD = 19,39) по сравнению с нейтральными целями ( M = 386,85, SD = 17,26). Никакие другие эффекты не были значительными [все F s (1100) < 3,44, нс , η 2 p < 0,04; Рисунок 6А]. Рисунок 6. Выполнение задачи в зависимости от типа цели, типа блока и группы в исследовании 2 . Отображается (A) время реакции в go-trials в мс, (B) количество совершенных ошибок и (C) пропуск ошибок. Столбики погрешностей указывают стандартные ошибки среднего. *** р < 0,001. Были основные эффекты для целевого типа [ F (1, 100) = 58,01, P <0,001, η 2 P 0 2 P 0 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341 = 0,341. F (1, 100) = 130,66, p < 0,001, η 2 p = 0,57], что указывает на то, что участники совершили больше ошибок в нейтральных блоках ( M = 10,20 ошибок, SD = 4,32) по сравнению с пищевыми блоками ( M = 6,92 ошибки, SD = 4,16) и в блоках сдвига ( M = 10,87 ошибок, SD = 4,87) по сравнению с блоками без сдвига ( M = 6,25 ошибок, SD = 3,34). Были основные эффекты для типа цели [ F (1, 100) = 94,83, P <0,001, η 2 P 9088 2 P 0 = 0,41 = 0,41 = 0,41 = 0,41 = 0,41 = 0,41 =. F (1, 100) = 68,46, p < 0,001, η 2 p = 0,41], что указывает на то, что испытуемые пропустили больше мишеней в нейтральных блоках ( M = 1,2 = 1,2 = 0,41).0887 SD = 6,86) по сравнению с пищевыми блоками ( M = 7,75 ошибок, SD = 5,39) и в сменных блоках ( M = 12,32 ошибок, SD = 6,93) по сравнению с несменными блоками ( М = 8,12 ошибок, SD = 5,30). Никакие другие эффекты не были значимыми [все F s (1100) < 2,69, нс , η 2 p < 0,03; Рисунок 6С]. Групповая, пищевая депривация и состояние тяги к еде вообще не предсказывают выполнение задания (табл. 5). Взаимодействие 9Группа 0887 × ИМТ предсказал ошибки пропусков в пищевых блоках: ИМТ положительно предсказывал ошибки пропусков в пищевых блоках в голодной группе (β = 0,34, p <0,05), но не в сытой группе (β = -0,16, нс ). Таблица 5. Прогнозирование выполнения задач в зависимости от пищевой депривации, состояния тяги к еде, ИМТ, успешности диеты, импульсивности и группы (исследование 2) . Успех диеты предсказывал более быстрое время реакции и меньшее количество ошибок упущения в нейтральных блоках и более медленное время реакции на пищу по сравнению с нейтральными блоками (таблица 5). Были и другие значимые взаимодействия 9Группа 0887 × успех диеты при прогнозировании выполнения задач в пищевых блоках (таблица 5): успех диеты был связан с более быстрым временем реакции (β = -0,28, p < 0,05), меньшим количеством комиссионных ошибок (β = -0,28, p <0,05) и ошибки пропусков (β = -0,51, p <0,001) у голодных участников, но не у сытых участников (все βs <0,24, нс ; рис. Рис. 7. Выполнение задания в блоках с пищевыми целевыми показателями в зависимости от группы и успеха диеты в исследовании 2: (A) время реакции в пробах в мс, (B) количество совершенных ошибок и (C) пропуск ошибок . Все переменные z -стандартизированы. В нейтральных блоках самооценка импульсивности была связана с большим количеством комиссионных ошибок (Таблица 5). Были и другие значимые взаимодействия группа × импульсивность при прогнозировании ошибок совершения: импульсивность была связана с большим количеством ошибок совершения в нейтральных блоках (β = 0,50, p < 0,001) и с меньшим количеством ошибок совершения в еде по сравнению с нейтральными блоками (β = -0,36 , p <0,01) у сытых участников, но не у голодных участников (βs <0,08, нс ). В исследовании 2 половина участников потребляла пищу перед выполнением задания, что успешно уменьшило их текущую тягу к еде на протяжении всего задания по сравнению с голодной группой. Тем не менее, состояние тяги к еде увеличилось после выполнения задания по сравнению с тем, что было до него, в обеих группах. Более высокая тяга к еде как до, так и после задания была связана с более низким ИМТ, и остается спекулятивным, если это можно объяснить физиологическими механизмами или предвзятостью сообщения, такой как социальная желательность. Повышение тяги к еде во время выполнения задания было предсказано более низким успехом диеты у сытых, но не у голодных участников. Это открытие может отражать эффект растормаживания предварительной нагрузки (Herman and Mack, 19). Повторяя результаты исследования 1, исследование 2 показало, что участники быстрее реагировали на пищу, чем нейтральные мишени, и пропускали меньше еды, чем нейтральные мишени. В отличие от исследования 1, участники также совершили меньше ошибок в пищевых блоках. Таким образом, результаты повторяют результаты, показывающие снижение поведенческого торможения в нейтральных блоках (т. е. более импульсивные реакции на пищевые дистракторы) в AST (Loeber et al., 2012, 2013). Различия всех трех показателей выполнения задания между пищевым и нейтральным блоками не зависели от приема пищи, т. е. не отличались между голодными и сытыми особями. Как и в исследовании 1, пищевая депривация и тяга к еде не были связаны с выполнением задания. Голодные участники пропустили больше пищевых целей с более высоким ИМТ и более низким успехом на диете, соответственно. Ассоциации не могли быть найдены у сытых участников, и, по сути, отношения, казалось, были обратными, поскольку сытые участники пропускали меньше еды, чем нейтральные целевые группы с более низким успехом диеты. Более высокий успех диеты также был связан с более быстрой реакцией и меньшим количеством ошибок у голодных, но не у сытых участников. Наконец, самооценка импульсивности была связана с большим количеством ошибок в нейтральных блоках только в сытой группе. Подводя итог, результаты показывают, что поведенческое торможение, вызванное пищевыми сигналами, не связано с текущим голодом или тягой, то есть, что различия в выполнении задачи между пищевыми и нейтральными стимулами не могут быть объяснены этими переменными. В текущих исследованиях мы исследовали влияние пищевых сигналов на поведенческое торможение с помощью графического AST. В исследовании 1 участники показали ускоренное время реакции в ответ на пищевые мишени по сравнению с нейтральными мишенями. Они также пропускали меньше еды, чем нейтральные цели. Эти результаты могут быть воспроизведены в исследовании 2 и согласуются с предыдущими исследованиями, в которых использовались лексические AST, связанные с едой (Mobbs et al. Другим объяснением различий во времени реакции и ошибках пропусков между пищевыми и нейтральными блоками может быть просто эффект размера категории (Landauer and Freedman, 1968). Изображения еды могут быть распознаны быстрее, чем нейтральные изображения, поскольку категория еды меньше, чем более широкая категория нейтральных объектов. Лобер и др. (2013) попытались решить эту проблему, используя слова одежды только в качестве контрольной категории. К сожалению, время реакции в этом исследовании не сообщалось. В заключение, нельзя полностью исключить возможный эффект размера категории, лежащий в основе различий в этих показателях выполнения задач. Результаты комиссионных ошибок различались между исследованиями 1 и 2. Ошибки комиссии были связаны с ИМТ и успехом диеты. В исследовании 1 результаты показали более выраженную расторможенность в еде по сравнению с нейтральными блоками с более низким успехом на диете и более высоким ИМТ. Эти результаты могут быть частично воспроизведены у голодных участников исследования 2, так что более низкий успех диеты был связан с усилением расторможенности в пищевых блоках. В недавнем исследовании Houben et al. (2012b), неудачники, сидящие на диете, также были менее подавлены в задаче стоп-сигнала, связанной с едой, после воздействия пищи, в то время как люди, успешно сидящие на диете, не пострадали. Наши результаты подтверждают эти выводы в том, что как успешные, так и неуспешные люди, сидящие на диете, могут быть в равной степени соблазнены вкусными пищевыми сигналами, но только у успешных людей, сидящих на диете, такие сигналы могут активировать механизм контроля (Stroebe et al. Хотя потребление пищи не влияло на выполнение задачи в целом, оно влияло на выполнение задачи в зависимости от успеха диеты. То есть связь между выполнением задачи и успехом на диете, которая была обнаружена у голодных участников, исчезла или даже изменилась на противоположную у сытых участников. Этот вывод может быть частично объяснен эффектом предварительной нагрузки-растормаживания (Herman and Mack, 1975). Точно так же, как известно, что неудачники, сидящие на диете, увеличивают потребление пищи после употребления предварительной нагрузки (Рудерман, 1986; Heatherton et al., 1988), низкий успех диеты был связан с более сильным увеличением тяги к еде во время выполнения задания после приема пищи. Однако текущая тяга к еде не была связана с выполнением задачи, поэтому изменения в выполнении задачи в результате приема пищи и успеха диеты не могут быть объяснены изменениями в текущей тяге к еде. Другим интересным открытием настоящих исследований было то, что самооценка импульсивности не была связана с поведенческим торможением у голодных участников, но у сытых участников можно было наблюдать взаимосвязь. То есть сытые участники совершали больше ошибок в нейтральных блоках с возрастающей импульсивностью, о которой они сообщали. Это открытие имеет важное значение для будущих исследований. Например, недавний метаанализ показал, что существует положительная связь между импульсивностью и предвзятостью внимания, связанной с веществами (Coskunpinar and Cyders, 2013). Хотя эта взаимосвязь не зависела от типа вещества (например, пищи или наркотиков), существовала более сильная связь между предвзятостью внимания и поведенческой импульсивностью (по оценке с помощью поведенческих задач), чем личностной импульсивностью (по оценке с помощью анкет). Похоже, что влияние пищевых сигналов на поведенческое торможение можно наблюдать только тогда, когда эти сигналы фактически используются в качестве целей и отвлекающих факторов, соответственно (Mobbs et al., 2008, 2011; Houben et al., 2012b). ; Loeber et al., 2012, 2013; Nederkoorn et al., 2012; Meule et al., 2014), но не тогда, когда используются нейтральные стимулы, например, письма, которые просто сопровождаются пищевыми сигналами (Meule et al. , 2011a, 2012c) или при воздействии пищевых продуктов (Nederkoorn et al., 2004). В отличие от предыдущих исследований, в которых использовался лексический AST, связанный с едой, мы использовали изображения еды, которые, возможно, имеют более высокую внешнюю валидность по сравнению со словами о еде и, как следствие, оказывают большее влияние на когнитивные задачи (Brooks et al., 2011). Однако ограничение текущего исследования заключается в том, что мы не использовали другую существенную категорию непродовольственных товаров (см. Что касается клинических последствий, недавние исследования показывают, что импульсивность или поведенческая расторможенность поддаются модификации (Guerrieri et al., 2009, 2012) и что обучение торможению снижает последующее потребление пищи (Houben, 2011; Houben and Jansen, 2011). Например, когда релевантные стимулы (например, изображения шоколада) сочетались с остановочной реакцией, снижение потребления закусок у лиц с низким тормозным контролем (Houben, 2011) и снижение потребления шоколада у любителей шоколада (Houben and Jansen, 2011) можно было наблюдать. Кроме того, было обнаружено, что постоянное сочетание вкусных пищевых сигналов со стоп-сигналами снижает потребление этих продуктов у хронически сидящих на диете (Veling et al. В заключение, настоящее исследование показало, что изобразительные, высококалорийные пищевые сигналы влияют на торможение двигательных реакций. Кроме того, выполнение задачи было по-разному связано с ИМТ, самооценкой успеха диеты и импульсивностью в зависимости от голода и насыщения. Тем не менее, мы исследовали выборку здоровых молодых женщин с нормальным ИМТ. Будущие исследования могут расширить текущие результаты и оценить их клиническую значимость путем изучения образцов с ожирением или людей с расстройствами пищевого поведения. Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов. Финансирование этого исследования было обеспечено грантом учебно-исследовательской группы 1253/2, который поддерживается Немецким исследовательским фондом (Deutsche Forschungsgemeinschaft, DFG) из федерального и земельного фондов. Адамс, С., Атайя, А.Ф., Аттвуд, А.С., и Мунафо, М.Р. (2013). Влияние алкоголя на растормаживание по отношению к сигналам, связанным с алкоголем. Наркотики Алкоголь Зависимость . 127, 137–142. doi: 10.1016/j.drugalcdep.2012.06.025 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Aichert, D.S., Wostmann, N.M., Costa, A., Macare, C., Wenig, J.R., Moller, H.J., et al. (2012). Связь между чертой импульсивности и доминантным торможением реакции. Дж. Клин. Эксп. Нейропсихология . 34, 1016–1032. doi: 10.1080/13803395.2012.706261 Опубликовано Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка Appelhans, BM (2009). Нейроповеденческое торможение кормления, основанного на вознаграждении: последствия для диеты и ожирения. Ожирение 17, 640–647. doi: 10.1038/oby.2008.638 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Аппельханс, Б. Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Баттеринк Л., Йокум С. и Стайс Э. (2010). Масса тела обратно пропорциональна тормозному контролю в ответ на еду у девочек-подростков: исследование фМРТ. Нейроизображение 52, 1696–1703. doi: 10.1016/j.neuroimage.2010.05.059 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Брукс, С., Принс, А., Шталь, Д., Кэмпбелл, И. К., и Треже, Дж. (2011). Систематический обзор и метаанализ когнитивной предвзятости к пищевым стимулам у людей с расстройствами пищевого поведения. клин. Психол. Версия . 31, 37–51. doi: 10.1016/j.cpr.2010.09.006 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Карнелл С. Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Сепеда-Бенито, А., Фернандес, М.С., и Морено, С. (2003). Взаимосвязь симптомов пола и расстройства пищевого поведения с сообщаемой тягой к еде: построить валидацию опросников о состоянии и чертах тяги на испанском языке. Аппетит 40, 47–54. doi: 10.1016/S0195-6663(02)00145-9 Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Сепеда-Бенито, А., Гливс, Д.Х., Уильямс, Т.Л., и Эрат, С.А. (2000). Разработка и валидация опросников состояний и черт пищевой тяги. Поведение. Номер . 31, 151–173. doi: 10.1016/S0005-7894(00)80009-X CrossRef Полный текст Claes, L., Mitchell, JE, and Vandereycken, W. (2012). Вне контроля? Процессы торможения при расстройствах пищевого поведения с личностной и когнитивной точек зрения. Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Claes, L., Nederkoorn, C., Vandereycken, W., Guerrieri, R., and Vertommen, H. (2006). Импульсивность и отсутствие тормозящего контроля при расстройствах пищевого поведения. Ешьте. Поведение . 7, 196–203. doi: 10.1016/j.eatbeh.2006.05.001 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Коэн, Д. А., и Фарли, Т. А. (2008). Еда как автоматическое поведение. Пред. Хронический Дис . 5, 1–7. Опубликован Аннотация | Полный текст в публикации Коскунпинар, А., и Сайдерс, М.А. (2013). Импульсивность и предвзятость внимания, связанная с веществами: метааналитический обзор. Наркотики Алкоголь Зависимость . 133, 1–14. doi: 10.1016/j.drugalcdep.2013.05.008 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Дельгадо-Рико, Э., Рио-Валле, Дж. Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Даймонд, А. (2013). Исполнительные функции. год. Преподобный Психол . 64, 135–168. doi: 10.1146/annurev-psych-113011-143750 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Федоров И.С., Поливи Дж. и Герман С.П. (1997). Влияние предварительного воздействия пищевых сигналов на пищевое поведение сдержанных и несдержанных едоков. Аппетит 28, 33–47. doi: 10.1006/appe.1996.0057 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Фишбах, А., Фридман, Р.С., и Круглански, А.В. (2003). Не подвести нас к искушению: сиюминутные соблазны вызывают первостепенную активацию цели. Дж. Перс. соц. Психол . Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Галимберти Э., Мартони Р. М., Каваллини М. К., Эрцеговеси С. и Беллоди Л. (2012). Моторное торможение и когнитивная гибкость при подтипах расстройств пищевого поведения. Прог. Нейропсихофармакол. биол. Психиатрия 36, 307–312. doi: 10.1016/j.pnpbp.2011.10.017 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Guerrieri, R., Nederkoorn, C., and Jansen, A. (2008). Влияние импульсивной личности на переедание и ожирение: современное состояние дел. Психология. Топ . 17, 265–286. Геррьери, Р., Недеркоорн, К., и Янсен, А. (2012). Растормаживанию легче научиться, чем торможению. Влияние тренировки (рас)торможения на прием пищи. Аппетит 59, 96–99. doi: 10.1016/j.appet.2012.04.006 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Герьери Р., Недеркоорн К., Шроутен М. Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Heatherton, T. F., Herman, C. P., Polivy, J., King, G. A., and McGree, S. T. (1988). (Неправильное) измерение сдержанности: анализ концептуальных и психометрических вопросов. Дж. Ненормальный. Психол . 97, 19–28. doi: 10.1037/0021-843X.97.1.19 Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Хендрик, О. М., Луо, X., Чжан, С., и Ли, К. С. Р. (2012). Обработка значимости и ожирение: предварительное исследование визуализации задачи стоп-сигнала. Ожирение 20, 1796–1802. doi: 10.1038/oby.2011.180 Pubmed Abstract | Опубликован полный текст | Полнотекстовая перекрестная ссылка Герман, С. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Хофманн В., Фризе М. и Рофс А. (2009). Три способа сопротивляться искушению: независимые вклады управляющего внимания, тормозного контроля и аффектной регуляции в импульсный контроль пищевого поведения. Дж. Экспл. соц. Психол . 45, 431–435. doi: 10.1016/j.jesp.2008.09.013 Полный текст CrossRef Хофманн В., Шмейхель Б. Дж. и Баддели А. Д. (2012). Исполнительные функции и саморегуляция. Познание тенденций. Наука . 16, 174–180. doi: 10.1016/j.tics.2012.01.006 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Хоу Р., Могг К., Брэдли Б. П., Мосс-Моррис Р., Певелер Р. и Рофс А. (2011). Внешнее переедание, импульсивность и склонность внимания к пищевым сигналам. Аппетит 56, 424–427. doi: 10.1016/j.appet.2011. Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef Houben, K. (2011). Преодоление желания разориться: влияние на пищевое поведение путем манипулирования тормозящим контролем. Дж. Бехав. тер. Эксп. Психиатрия 42, 384–388. doi: 10.1016/j.jbtep.2011.02.008 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Houben, K., Havermans, R.C., Nederkoorn, C., and Jansen, A. (2012a). Пиво запрещено: обучение прекращению реакции на алкогольные сигналы снижает потребление алкоголя за счет уменьшения аффективных ассоциаций, а не усиления торможения реакции. Дополнение 107, 1280–1287. doi: 10.1111/j.1360-0443.2012.03827.x Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Хоубен, К., и Янсен, А. (2011). Тренировка тормозного контроля. Рецепт сопротивления сладким искушениям. Аппетит 56, 345–349. doi: 10.1016/j.appet.2010.12.017 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Houben, K. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Янсен, А., Недеркоорн, К., ван Баак, Л., Кейрс, К., Гуерьери, Р., и Хаверманс, Р. (2009). Те, кто сдержан в еде, переедают только тогда, когда они импульсивны. Поведение. Рез. Номер . 47, 105–110. doi: 10.1016/j.brat.2008.10.016 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Jansen, A., Theunissen, N., Slechten, K., Nederkoorn, C., Boon, B., Mulkens, S., et al. (2003). Дети с избыточным весом переедают после воздействия пищевых сигналов. Ешьте. Поведение . 4, 197–209. doi: 10.1016/S1471-0153(03)00011-4 Опубликовано Аннотация | Опубликован полный текст | CrossRef Полный текст Ясинска, А. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Кемпс, Э., и Тиггеманн, М. (2010). Когнитивный экспериментальный подход к пониманию и уменьшению тяги к еде. Курс. Реж. Психол. Наука . 19, 86–90. doi: 10.1177/0963721410364494 CrossRef Full Text Landauer, T.K., and Freedman, J.L. (1968). Извлечение информации из долговременной памяти: размер категории и время распознавания. J. Глагол. Учиться. Глагол. Поведение . 7, 291–295. doi: 10.1016/S0022-5371(68)80003-9 CrossRef Полный текст Лебер С., Гроссханс М., Херперц С., Кифер Ф. и Герперц С. (2013). Голод модулирует поведенческую расторможенность и распределение внимания на сигналы, связанные с едой, у контрольных людей с нормальным весом. Pubmed Abstract | Опубликован полный текст | CrossRef Полный текст Лебер, С., Гроссханс, М., Корукуоглу, О., Фоллмерт, К., Фоллштадт-Кляйн, С., Шнайдер, С., и др. (2012). Нарушение тормозного контроля в ответ на сигналы, связанные с едой, и предвзятость внимания участников с ожирением и контрольной группы с нормальным весом. Междунар. Дж. Обес . 36, 1334–1339. doi: 10.1038/ijo.2011.184 Pubmed Abstract | Опубликован полный текст | Полнотекстовая перекрестная ссылка Логан, Г. Д., Шачар, Р. Дж., и Таннок, Р. (1997). Импульсивность и тормозящий контроль. Психология. Наука . 8, 60–64. doi: 10.1111/j.1467-9280.1997.tb00545.x Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Lowe, MR (2003). Саморегуляция потребления энергии в профилактике и лечении ожирения: возможно ли это? Обес. Рез . 11, 44с–59с. doi: 10.1038/oby.2003.223 Опубликовано Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка Лоу, М. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Лоу, М. Р., и Бутрин, М. Л. (2007). Гедонистический голод: новое измерение аппетита? Физиол. Поведение . 91, 432–439. doi: 10.1016/j.physbeh.2007.04.006 Pubmed Abstract | Опубликован полный текст | Полнотекстовая перекрестная ссылка Меуле, А. (2013). Импульсивность и переедание: более пристальный взгляд на подшкалы шкалы импульсивности Барратта. Фронт. Психол . 4:177. doi: 10.3389/fpsyg.2013.00177 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Меул, А., и Блехерт, Дж. (2012). food.pics: база данных изображений для изучения питания и аппетита. Обес. Facts 5, 20. doi: 10.1159/000170914 CrossRef Full Text Meule, A., Lukito, S. Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Меуле, А., Лутц, А., Фёгеле, К., и Кюблер, А. (2012a). Тяга к еде по-разному различает успешных и неудачников, сидящих на диете, и тех, кто не сидит на диете. Валидация опросников о тяге к еде на немецком языке. Аппетит 58, 88–97. doi: 10.1016/j.appet.2011.09.010 Pubmed Abstract | Опубликован полный текст | Полнотекстовая перекрестная ссылка Меуле, А., Лутц, А., Фёгеле, К., и Кюблер, А. (2012b). Самооценка успеха диеты связана с вегетативной регуляцией сердца у тех, кто сидит на диете. Аппетит 59, 494–498. doi: 10.1016/j.appet.2012.06.013 Pubmed Abstract | Опубликован полный текст | CrossRef Полный текст Меуле, А., Лутц, А., Фёгеле, К., и Кюблер, А. (2012c). Женщины с повышенными симптомами пищевой зависимости демонстрируют ускорение реакции, но не нарушение тормозного контроля, в ответ на изображения высококалорийных пищевых сигналов. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Меуле, А., Лутц, А., Фёгеле, К., и Кюблер, А. (2014). Импульсивные реакции на пищевые сигналы предсказывают последующую тягу к еде. Ешьте. Поведение . 15, 99–105. doi: 10.1016/j.eatbeh.2013.10.023 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Меуле, А., Папиес, Е.К., и Кюблер, А. (2012d). Дифференциация между успешными и неудачливыми людьми, сидящими на диете: достоверность и надежность шкалы воспринятого саморегулирующегося успеха в диете. Аппетит 58, 822–826. doi: 10.1016/j.appet.2012.01.028 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Меуле, А., Скирде, А.К., Фройнд, Р., Фёгеле, К., и Кюблер, А. (2012e). Высококалорийные пищевые сигналы ухудшают работу рабочей памяти у людей с высокой и низкой тягой к еде. Аппетит 59, 264–269. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Меуле, А., Фёгеле, К., и Кюблер, А. (2011b). [Психометрическая оценка немецкой шкалы импульсивности Барратта — краткая версия (BIS-15)]. Диагностика 57, 126–133. doi: 10.1026/0012-1924/a000042 CrossRef Full Text Meule, A., Westenhöfer, J., and Kübler, A. (2011c). Пристрастия к еде опосредуют связь между жестким, но не гибким контролем пищевого поведения и успехом диеты. Аппетит 57, 582–584. doi: 10.1016/j.appet.2011.07.013 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Моббс О., Иглесиас К., Голей А. и Ван дер Линден М. (2011). Когнитивный дефицит у людей с ожирением с компульсивным перееданием и без него. Исследование с использованием задачи на гибкость мышления. Аппетит 57, 263–271. doi: 10.1016/j.appet.2011.04.023 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Моббс, О. Pubmed Abstract | Опубликован полный текст | CrossRef Полный текст Мерфи, Ф. К., Саакян, Б. Дж., Рубинштейн, Дж. С., Майкл, А., Роджерс, Р. Д., Роббинс, Т. В., и др. (1999). Эмоциональная предвзятость и процессы тормозного контроля при мании и депрессии. Психология. Мед . 29, 1307–1321. doi: 10.1017/S003329179 33 Опубликовано Аннотация | Опубликован полный текст | CrossRef Full Text Nederkoorn, C., Braet, C., Van Eijs, Y., Tanghe, A., and Jansen, A. (2006a). Почему дети с ожирением не могут устоять перед едой: роль импульсивности. Ешьте. Поведение . 7, 315–322. doi: 10.1016/j.eatbeh.2005.11.005 Pubmed Abstract | Опубликован полный текст | Полнотекстовая перекрестная ссылка Nederkoorn, C. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Недеркоорн, К., Хоубен, К., Хофманн, В., Руфс, А., и Янсен, А. (2010). Контролировать себя или просто есть то, что нравится? Прибавка в весе в течение года прогнозируется интерактивным эффектом торможения реакции и неявным предпочтением закусок. Психология здоровья . 29, 389–393. doi: 10.1037/a0019921 Опубликовано Аннотация | Опубликован полный текст | CrossRef Full Text Nederkoorn, C., Jansen, E., Mulkens, S., and Jansen, A. (2007). Импульсивность предсказывает исход лечения у детей с ожирением. Поведение. Рез. Номер . 45, 1071–1075. doi: 10.1016/j.brat.2006.05.009 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Nederkoorn, C., Smulders, F. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Nederkoorn, C., Van Eijs, Y., and Jansen, A. (2004). Сдержанные едоки действуют импульсивно. чел. Индивид. Отличие . 37, 1651–1658. doi: 10.1016/j.paid.2004.02.020 Полный текст CrossRef Nigg, JT (2000). О торможении / расторможенности в психопатологии развития: взгляды когнитивной и личностной психологии и рабочая таксономия торможения. Психология. Бык . 126, 220–246. doi: 10.1037/0033-2909.126.2.220 Опубликовано Аннотация | Опубликован полный текст | CrossRef Full Text Noël, X., Van der Linden, M., d’Acremont, M., Bechara, A., Dan, B., Hanak, C., et al. (2007). Алкогольные сигналы усиливают когнитивную импульсивность у людей, страдающих алкоголизмом. Психофармакол . 192, 291–298. doi: 10.1007/s00213-006-0695-6 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Noël, X. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Паттон, Дж. Х., Стэнфорд, М. С., и Барратт, Э. С. (1995). Факторная структура шкалы импульсивности Барратта. Дж. Клин. Психол . 51, 768–774. Опубликован Аннотация | Полный текст в публикации Росвал, Л., Стайгер, Х., Брюс, К., Исраэль, М., Ричардсон, Дж., и Обут, М. (2006). Импульсивность у женщин с расстройствами пищевого поведения: проблема торможения реакции, планирования или внимания? Междунар. Дж. Ешьте. Беспорядок . 39, 590–593. doi: 10.1002/eat.20296 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Ruderman, AJ (1986). Диетическое ограничение: теоретический и эмпирический обзор. Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Шульц К.П., Фан Дж., Магидина О., Маркс Д.Дж., Хан Б. и Гальперин Дж.М. (2007). Действительно ли эмоциональная задача «да/нет» измеряет поведенческую заторможенность? Сходимость с мерами по неэмоциональному аналогу. Арх. клин. Нейропсихология . 22, 151–160. doi: 10.1016/j.acn.2006.12.001 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Спинелла, М. (2007). Нормативные данные и краткая форма шкалы импульсивности Барратта. Междунар. Дж. Нейроски . 117, 359–368. doi: 10.1080/00207450600588881 Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Stroebe, W. (2008). «Ограниченное питание и нарушение саморегуляции», в Диета, избыточный вес и ожирение — саморегуляция в среде, богатой пищевыми продуктами г., изд. В. Стробе (Вашингтон, округ Колумбия: Американская психологическая ассоциация), 115–139. Опубликовано Аннотация | Опубликован полный текст | CrossRef Full Text Stroebe, W., Van Koningsbruggen, G.M., Papies, E.K., and Aarts, H. (2013). Почему большинство людей, сидящих на диете, терпят неудачу, но некоторые добиваются успеха: модель пищевого поведения с конфликтом целей. Психология. Версия . 120, 110–138. doi: 10.1037/a0030849 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Тетли, А.С., Бранстром, Дж.М., и Гриффитс, П.Л. (2010). Роль чувствительности к вознаграждению и импульсивности в реакции на пищевой сигнал. Ешьте. Поведение . 11, 138–143. doi: 10.1016/j.eatbeh.2009.12.004 Pubmed Abstract | Опубликован полный текст | CrossRef Полный текст Ван ден Эйнде, Ф., Самаравикрема, Н., Кеньон, М., Де Йонг, Х., Лавендер, А., Стартап, Х., и др. (2012). Исследование нейрокогнитивных функций при нервной булимии и расстройстве пищевого поведения, не указанном иначе, — тип булимии. Опубликовано Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка Вандер Вал, Дж. С., Джонстон, К. А., и Дурандхар, Н. В. (2007). Психометрические свойства опросников State and Trait Food Cravings у лиц с избыточной массой тела и ожирением. Ешьте. Поведение . 8, 211–223. doi: 10.1016/j.eatbeh.2006.06.002 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Van Koningsbruggen, G.M., Stroebe, W., and Aarts, H. (2013). Взлет и падение самоконтроля: активация цели, вызванная искушением, и целенаправленное поведение, требующее усилий. Соц. Психол. Перс. Наука . 4, 546–554. doi: 10.1177/1948550612471061 CrossRef Full Text Van Koningsbruggen, G.M., Veling, H., Stroebe, W. и Aarts, H. (в печати). Сравнение двух психологических вмешательств по уменьшению импульсивных процессов пищевого поведения: влияние на самостоятельно выбранный размер порции. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Veling, H., Aarts, H., and Papies, E.K. (2011). Использование стоп-сигналов для подавления реакции людей, сидящих на диете, на вкусную пищу. Поведение. Рез. Номер . 49, 771–780. doi: 10.1016/j.brat.2011.08.005 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Велинг Х., Аартс Х. и Стробе В. (2013a). Использование стоп-сигналов для снижения импульсивного выбора вкусной нездоровой пищи. Бр. J. Health Psychol . 18, 354–368. doi: 10.1111/j.2044-8287.2012.02092.x Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Велинг Х., Аартс Х. и Стробе В. (2013b). Стоп-сигналы уменьшают выбор вкусных продуктов за счет снижения оценки продуктов. Фронт. Психол . 4:875. doi: 10.3389/fpsyg.2013.00875 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Westenhoefer, J. Pubmed Abstract | Опубликован полный текст | CrossRef Полный текст Ву, М., Гиль, К. Э., Скунде, М., Шаг, К., Рудофски, Г., де Цваан, М., и др. (2013). Тормозной контроль и принятие решений в условиях риска при нервной булимии и компульсивном переедании. Междунар. Дж. Ешьте. Беспорядок . 46, 721–728. doi: 10.1002/eat.22143 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Космокаликс зрелищный Этот вид происходит с полуострова Юкатан в Мексике. Его также называют Красносердым. В свежесрезанном виде он ярко-красного цвета, который темнеет при выдержке. Номер предмета:
W40038 В40038А $7,25 А 1 9/16 х 1 9/16 x 9 7/16″ W40038B 6,55 долларов США B 1 1/2 x 1 1/2 x 9 1/4 дюйма W40038C $6,95 C 1 1/2 x 1 1/2 x 9 7/8″ W40038D $8,70 D 1 1/2 x 1 1/2 x 12 5/16″ W40038E $9,30 E 1 9/16 x 1 9/16 x 12 1/8″ W40038F $8,80 F 1 1/2 x 1 1/2 x 12 1/2 дюйма W40038G $9,65 Г 1 9/16 х 1 9/16 x 12 5/8″ W40038H $9,35 В 1 1/2 x 1 1/2 x 13 1/4 дюйма Номер предмета:
W40018 В40018А $18. A 1 3/4 x 1 3/4 x 25 3/16 дюйма $18.00 B 1 5/8 x 1 5/8 x 30 1/8″ W40018C 20 долларов США C 1 3/4 x 1 3/4 x 30 1/8″ W40018D 20 долларов США D 1 3/4 x 1 3/4 x 30 1/16 дюйма W40018E 20 долларов США E 1 11/16 x 1 11/16 x 30 1/8″ W40018F 20 долларов США F 1 3/4 x 1 3/4 x 30 1/8″ W40018G 20 долларов США G 1 11/16 x 1 11/16 x 30 1/8″ W40018H $18.00 В 1 3/4 x 1 3/4 x 29 1/16 дюйма Хлороксилон свиетения Эта древесина поступает в основном из Шри-Ланки и уже несколько столетий является ценным товарным деревом. Дерево выглядит так, что к нему хочется прикоснуться. Номер предмета:
W40014 В40014А $30.00 A 1 3/4 x 1 3/4 x 21 15/16″ W40014B $35.00 B 1 11/16 x 1 11/16 x 24 дюйма W40014C $35.00 C 1 3/4 x 1 3/4 x 24 дюйма W40014D $35.00 D 1 3/4 x 1 3/4 x 24 1/16 дюйма W40014E $35.00 Е 1 13/16 х 1 13/16 х 25 дюймов Хлороксилон свиетения Эта древесина поступает в основном из Шри-Ланки и уже несколько столетий является ценной древесиной для торговли. Дерево выглядит так, что к нему хочется прикоснуться. После тонкой полировки и отделки Satinwood имеет гладкую, шелковистую поверхность с блестящей глубиной. Номер предмета:
W40000 24,40 доллара США A 1 9/16 x 1 9/16 x 22 1/8″ W40000B 24,50 доллара США B 1 9/16 x 1 9/16 x 22 3/16 дюйма 24,40 доллара США С 1 9/16 х 1 9/16 х 22 1/8 дюйма В40000Д 24,40 доллара США D 1 9/16 x 1 9/16 x 22 1/8″ W40000E 24,40 доллара США E 1 9/16 x 1 9/16 x 22 1/8″ W40000F 26,40 долларов США F 1 5/8 x 1 5/8 x 22 1/8″ W40000G 27,45 долларов США G 1 5/8 x 1 5/8 x 23 дюйма $31,65 В 1 3/4 x 1 3/4 x 22 7/8 дюйма Космокаликс зрелищный Этот вид происходит с полуострова Юкатан в Мексике. Номер предмета:
W39990 $11,30 A 1 9/16 x 1 9/16 x 14 3/4 дюйма W39990B $10,65 B 1 1/2 x 1 1/2 x 15 1/8″ 11,90 долларов США C 1 1/2 x 1 1/2 x 16 7/8 дюйма W39990D $13,30 D 1 9/16 x 1 9/16 x 17 3/8″ W39990E $16.00 E 1 1/2 x 1 1/2 x 22 3/4 дюйма $16,70 F 1 9/16 x 1 9/16 x 21 7/8″ Прозопис черный Очень немногие кусочки мескитового дерева украшены тем, что мы называем завитком лягушачьего волоса, маленьким, тугим завитком с 12-15 завитками на дюйм. Очень устойчивая древесина. Высушенный на воздухе. Номер предмета:
W39946 В39946А 22,95 доллара США A 1 9/16 x 1 9/16 x 19 7/8″ W39946B 22,85 доллара США B 1 9/16 x 1 9/16 x 19 13/16″ 21,15 доллара США C 1 1/2 x 1 1/2 x 19 7/8 дюйма W39946D 21,10 доллара США D 1 1/2 x 1 1/2 x 19 13/16″ 22,85 доллара США E 1 9/16 x 1 9/16 x 19 13/16″ W39946F 22,85 доллара США F 1 9/16 x 1 9/16 x 19 13/16″ 22,95 доллара США G 1 9/16 x 1 9/16 x 19 7/8″ 22,95 доллара США В 1 9/16 x 1 9/16 x 19 7/8″ Прозопис черный Очень немногие кусочки мескитового дерева украшены тем, что мы называем завитком лягушачьего волоса, маленьким, тугим завитком с 12-15 завитками на дюйм. Изделие F с изысканным рисунком и одним из самых красивых изделий из фигурного мескитового дерева, которые мы когда-либо видели. Номер предмета:
W39945 В39945А 25 долларов США A 1 5/8 x 1 5/8 x 20 3/16 дюйма W39945B 16,85 долларов США B 1 7/16 x 1 7/16 x 21 5/16 дюйма W39945C 22,45 доллара США C 1 7/16 x 1 7/16 x 23 дюйма W39945D $22.00 D 1 1/2 x 1 1/2 x 25 9/16 дюйма W39945Э 21,60 доллара США E 1 1/2 x 1 1/2 x 25 1/8 дюйма $50.00 F 1 1/2 x 1 1/2 x 25 1/16 дюйма Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Номер предмета:
W39750 27,95 долларов США A 1 1/2 x 1 1/2 x 13 1/4 дюйма $31,20 B 1 9/16 x 1 9/16 x 13 5/8″ 26,65 долларов США C 1 7/16 x 1 7/16 x 13 3/4 дюйма 28,35 долларов США D 1 7/16 x 1 7/16 x 14 5/8″ 44,25 доллара США E 1 9/16 x 1 9/16 x 19 5/16 дюйма 49,65 долларов США F 1 9/16 x 1 9/16 x 21 11/16″ W39750G $52,40 G 1 9/16 x 1 9/16 x 22 7/8 дюйма $54,95 В 1 9/16 x 1 9/16 x 24 дюйма Диоспирос островной Эта древесина происходит с небольшого малонаселенного острова у восточного побережья Новой Гвинеи. Номер предмета:
W39442 $ A продано x x W39442B 30,75 долларов США B 15/16 x 3 3/4 x 14 13/16″ W39442C 22,10 доллара США C 15/16 x 2 9/16 x 15 9/16 дюйма 39,95 долларов США D 7/8 x 2 1/2 x 30 15/16″ 42,30 доллара США E 7/8 x 2 9/16 x 31 15/16″ Птерокарпус индикус Один из самых редких и красивых капов в мире. От светлого до глубокого красновато-оранжевого, иногда с более темными линиями, с прекрасным пряным ароматом, едва ли найдется участок, который не был бы ярко выражен с высоты птичьего полета. Номер предмета:
W39173 W39173A 225,00 долларов США A 1 1/2 x 1 1/2 x 18 1/16 дюйма 250,00 долларов США В 1 1/2 х 1 1/2 х 19 7/16 дюйма Диоспирос островной Эта древесина происходит с небольшого малонаселенного острова у восточного побережья Новой Гвинеи. Некоторые из этих деревьев вырастают до огромных размеров. Одно из первых бревен, которые мы получили отсюда, имело 3 1/2 фута в диаметре, 18 футов в длину и весило 7 тонн. Он мелкозернистый и имеет гладкую поверхность. Большинство этих изделий имеют вид «мраморного пирога» с выдающимся конечным зерном. Номер предмета:
W39122 17,50 долларов США A 1 9/16 x 1 9/16 x 13 3/4 дюйма $40.00 B 1 11/16 x 1 11/16 x 20 5/16″ W39122C 25,70 долларов США C 1 9/16 x 1 9/16 x 20 3/16 дюйма 29,95 долларов США D 1 11/16 x 1 11/16 x 20 3/16 дюйма W39122E $32,30 E 1 3/4 x 1 3/4 x 20 1/4 дюйма W39122F 25,60 долларов США F 1 9/16 x 1 9/16 x 20 1/8″ W39122Г 29,95 долларов США G 1 11/16 x 1 11/16 x 20 3/16 дюйма 30,05 долларов США В 1 11/16 x 1 11/16 x 20 1/4″ Гибуртия сп. Также называется Овангколь и Амазакуэ. Это дерево становится все более популярным в качестве дерева для акустических гитар. Цвет варьируется от насыщенного золотисто-коричневого до блестящего серовато-коричневого, обычно с черной полосой. Номер предмета:
W38947 $8,85 А 1 3/4 х 1 3/4 х 11 7/8 дюйма $11,30 B 1 3/4 x 1 3/4 x 15 1/8″ $13,45 C 1 3/4 x 1 3/4 x 18 1/16 дюйма W38947D $13,40 D 1 3/4 x 1 3/4 x 17 15/16″ W38947E 15,70 долларов США E 1 3/4 x 1 3/4 x 21 1/16 дюйма 15,70 долларов США F 1 3/4 x 1 3/4 x 21 1/16 дюйма 15,70 долларов США G 1 3/4 x 1 3/4 x 21 1/16 дюйма Свартзия фистулоидная Твердый и тяжелый, способный прекрасно полироваться. Номер предмета:
W38770 $ Продано x x 24,55 доллара США B 1 1/2 x 1 1/2 x 24 1/8 дюйма 25,60 долларов США C 1 9/16 x 1 9/16 x 23 3/16 дюйма W38770D 24,55 доллара США D 1 1/2 x 1 1/2 x 24 1/8 дюйма 24,55 доллара США E 1 1/2 x 1 1/2 x 24 1/8 дюйма 24,55 доллара США F 1 1/2 x 1 1/2 x 24 1/8″ Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Номер предмета:
W38651 $55.00 A 1 7/16 x 1 7/16 x 28 5/16 дюйма W38651B $57.00 B 1 7/16 x 1 7/16 x 29 13/16″ W38651C $60.00 C 1 1/2 x 1 1/2 x 31 3/16 дюйма $60.00 D 1 7/16 x 1 7/16 x 31 3/4 дюйма $60.00 E 1 7/16 x 1 7/16 x 31 7/8″ $60.00 F 1 7/16 x 1 7/16 x 31 11/16 дюйма $60. G 1 7/16 x 1 7/16 x 31 11/16 дюйма $60.00 В 1 7/16 x 1 7/16 x 31 11/16 дюйма Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. Номер предмета:
W38650 $60. A 1 1/2 x 1 1/2 x 31 11/16 дюйма $60.00 B 1 7/16 x 1 7/16 x 31 11/16 дюйма $ C продано x x W38650D $60.00 D 1 3/8 x 1 3/8 x 31 1/2 дюйма W38650E $60.00 E 1 1/2 x 1 1/2 x 31 13/16 дюйма $60.00 F 1 7/16 x 1 7/16 x 31 3/4 дюйма $60.00 G 1 7/16 x 1 7/16 x 31 3/4 дюйма Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Номер предмета:
W38639 $60.00 A 1 7/16 x 1 7/16 x 31 1/2 дюйма $55.00 B 1 5/16 x 1 5/16 x 31 11/16 дюйма $ C продано x x $60.00 D 1 7/16 x 1 7/16 x 31 9/16 дюйма W38639E $60.00 E 1 7/16 x 1 7/16 x 31 3/4 дюйма $60.00 F 1 7/16 x 1 7/16 x 31 9/16 дюйма W38639G $60. G 1 3/8 x 1 3/8 x 31 3/4 дюйма Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. Номер предмета:
W38173 W38173A $13,35 A 1 7/16 x 1 7/16 x 7 7/16 дюйма W38173B $13,60 B 1 7/16 x 1 7/16 x 7 9/16 дюйма W38173C 15,95 долларов США C 1 7/16 x 1 7/16 x 8 7/8 дюйма 20,95 долларов США D 1 3/8 x 1 3/8 x 12 3/4 дюйма 22,35 доллара США E 1 7/16 x 1 7/16 x 12 7/16 дюйма 27,85 долларов США F 1 7/16 x 1 7/16 x 15 1/2 дюйма $30,30 G 1 7/16 x 1 7/16 x 16 7/8″ Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Номер предмета:
W38171 $31,50 А 1 3/8 х 1 3/8 х 19 3/16 дюйма $36,45 B 1 7/16 x 1 7/16 x 20 5/16 дюйма 31,45 доллара США C 1 5/16 x 1 5/16 x 21 дюйм $34,10 D 1 3/8 x 1 3/8 x 20 3/4 дюйма $37.00 E 1 7/16 x 1 7/16 x 20 5/8″ W38171F $34,30 F 1 3/8 x 1 3/8 x 20 7/8 дюйма 41,30 доллара США G 1 1/2 x 1 1/2 x 21 1/8 дюйма $38,60 В 1 7/16 x 1 7/16 x 21 1/2 дюйма Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. На F один угол имеет обзол 5 дюймов на одном конце. Должен выключиться. Цена соответственно. Номер предмета:
W38170 В38170А 33,55 доллара США А 1 3/8 х 1 3/8 х 20 7/16 дюйма $36,80 B 1 7/16 x 1 7/16 x 20 1/2 дюйма 32,20 доллара США C 1 5/16 x 1 5/16 x 21 1/2 дюйма $35,10 D 1 3/8 x 1 3/8 x 21 3/8 дюйма $37,70 Е 1 7/16 х 1 7/16 х 21 дюйм $28. F 1 3/8 x 1 3/8 x 21 1/4 дюйма 30,70 долларов США G 1 5/16 x 1 5/16 x 20 1/2 дюйма $34,70 В 1 3/8 x 1 3/8 x 21 1/8″ Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. Номер предмета:
W37999 18,80 долларов США A 1 1/2 x 1 1/2 x 9 5/8 дюйма $33,70 B 1 1/2 x 1 1/2 x 17 1/4 дюйма W37999C 29,75 долларов США C 1 3/8 x 1 7/16 x 17 5/16 дюйма 27,75 долларов США D 1 1/4 x 1 1/4 x 20 7/16 дюйма $34,70 Е 1 3/8 х 1 3/8 х 21 1/8 дюйма Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Номер предмета:
W37998 $34,10 A 1 3/8 x 1 3/8 x 20 3/4 дюйма $37,45 B 1 7/16 x 1 7/16 x 20 7/8″ W37998C $34,40 C 1 3/8 x 1 3/8 x 20 15/16″ W37998D $35.00 D 1 3/8 x 1 3/8 x 21 5/16 дюйма $34,90 E 1 3/8 x 1 3/8 x 21 1/4 дюйма 35,50 долларов США F 1 3/8 x 1 3/8 x 21 5/8 дюйма Дальбергия барони Один из нескольких видов розового дерева с Мадагаскара. Поскольку этот товар находится в списке СИТЕС, мы отправляем этот вид только по адресам в США. Номер предмета:
W37587 $ Продано х х W37587B $52.00 B 1 1/4 x 1 1/4 x 33 7/16 дюйма W37587C $52.00 С 1 1/8 х 1 1/8 х 33 7/16 дюйма Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Номер предмета:
W36966 30,95 долларов США A 1 7/16 x 1 7/16 x 14 3/8 дюйма $32,60 B 1 7/16 x 1 7/16 x 15 1/8″ 35,95 долларов США C 1 7/16 x 1 7/16 x 16 11/16 дюйма 41,20 доллара США D 1 7/16 x 1 7/16 x 19 1/8″ W36966E $37,70 Е 1 3/8 х 1 3/8 х 19 1/8 дюйма 40,90 долларов США F 1 7/16 x 1 7/16 x 19 дюймов Диоспирос крассифлора Иногда его называют Габоном или Габуном, это черное дерево иногда бывает угольно-черным, но часто имеет сероватые полосы, которые в основном исчезают после завершения или после того, как изготовленный объект был введен в эксплуатацию. Номер предмета:
W36965 $44.00 A 1 1/2 x 1 1/2 x 18 3/4 дюйма $44.00 B 1 7/16 x 1 7/16 x 19 дюймов W36965C $44.00 С 1 7/16 х 1 7/16 х 19 1/16 дюйма Номер предмета:
W36635 W36635A $8,75 A 1 5/8 x 1 5/8 x 11 7/8 дюйма $8,75 B 1 1/2 x 1 1/2 x 14 дюймов $11,45 С 1 5/8 х 1 5/8 х 15 9/16 дюйма $14,75 D 1 5/8 x 1 5/8 x 20 1/16 дюйма $12,60 E 1 1/2 x 1 1/2 x 20 1/8 дюйма $14,80 F 1 5/8 x 1 5/8 x 20 1/8 дюйма 21,10 доллара США G 1 15/16 x 1 15/16 x 20 3/16 дюйма Рупала бразильская Красивое, блестящее зерно с большими серебристыми сердцевинными лучами. Номер предмета:
W36613 $17,10 А 1 7/8 х 1 7/8 х 20 дюймов 18,50 долларов США B 1 15/16 x 1 15/16 x 20 1/4″ W36613C $18,45 C 1 15/16 x 1 15/16 x 20 3/16 дюйма $18,45 D 1 15/16 x 1 15/16 x 20 3/16 дюйма 19,65 долларов США E 2 x 2 x 20 3/16 дюйма 20,15 долларов США F 2 x 2 x 20 11/16″ 20,15 долларов США G 2 x 2 x 20 11/16″ Энтандрофрагма цилиндрическая Дерево Сапеле растет в тропиках Западной и Центральной Африки. Даже без рисунка, как эта доска Sapele, имеет удивительный блеск и красиво полируется. Номер предмета:
W36335 В36335А 22,40 доллара США A 1 11/16 x 1 11/16 x 20 9/16 дюйма W36335B 22,40 доллара США B 1 11/16 x 1 11/16 x 20 9/16 дюйма 22,40 доллара США C 1 11/16 x 1 11/16 x 20 9/16″ W36335D 22,40 доллара США D 1 11/16 x 1 11/16 x 20 9/16 дюйма 22,40 доллара США E 1 11/16 x 1 11/16 x 20 9/16 дюйма 22,40 доллара США F 1 11/16 x 1 11/16 x 20 9/16 дюйма Гибуртия сп. Также называется Овангколь и Амазакуэ. Это дерево становится все более популярным в качестве дерева для акустических гитар. Цвет варьируется от насыщенного золотисто-коричневого до блестящего серовато-коричневого, обычно с черной полосой. Номер предмета:
W34268 $30,85 A 1 11/16 x 1 11/16 x 24 дюйма $30,85 B 1 11/16 x 1 11/16 x 24 дюйма $33,20 C 1 3/4 x 1 3/4 x 24 дюйма $33,35
26,68 долларов США D 1 3/4 x 1 3/4 x 24 1/8″ W34268E $33,35
26,68 долларов США E 1 3/4 x 1 3/4 x 24 1/8 дюйма $33,35 F 1 3/4 x 1 3/4 x 24 1/8″ Дальбергия меланоксилон Цвет сердцевины от лилового до коричневато-черного с темно-серыми прожилками, создающими общее впечатление черного цвета. ( М = 44,07, СО = 10,46), чем в группе сытости ( М = 31,63, СО = 10,11), и тяга была выше после выполнения задачи ( М СО = 40,887,
( М = 44,07, СО = 10,46), чем в группе сытости ( М = 31,63, СО = 10,11), и тяга была выше после выполнения задачи ( М СО = 40,887, Корреляты текущей тяги к еде
 5).
5). Производительность задачи
Время реакции
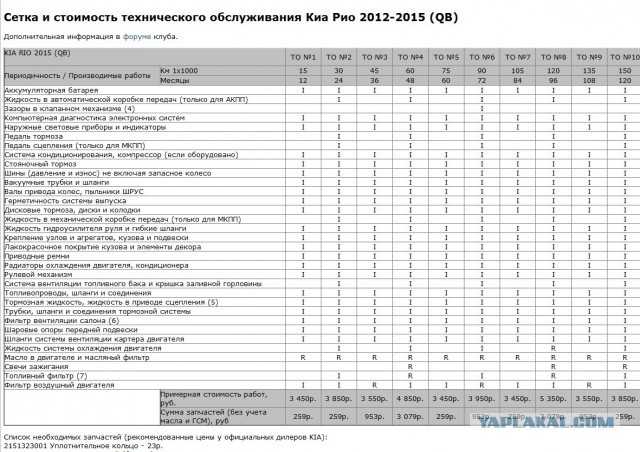
Комиссионные ошибки
 Никакие другие эффекты не были значительными [все F с (1,100) < 1,28, нс , η 2 р < 0,02; Рисунок 6Б].
Никакие другие эффекты не были значительными [все F с (1,100) < 1,28, нс , η 2 р < 0,02; Рисунок 6Б]. Ошибки пропущения
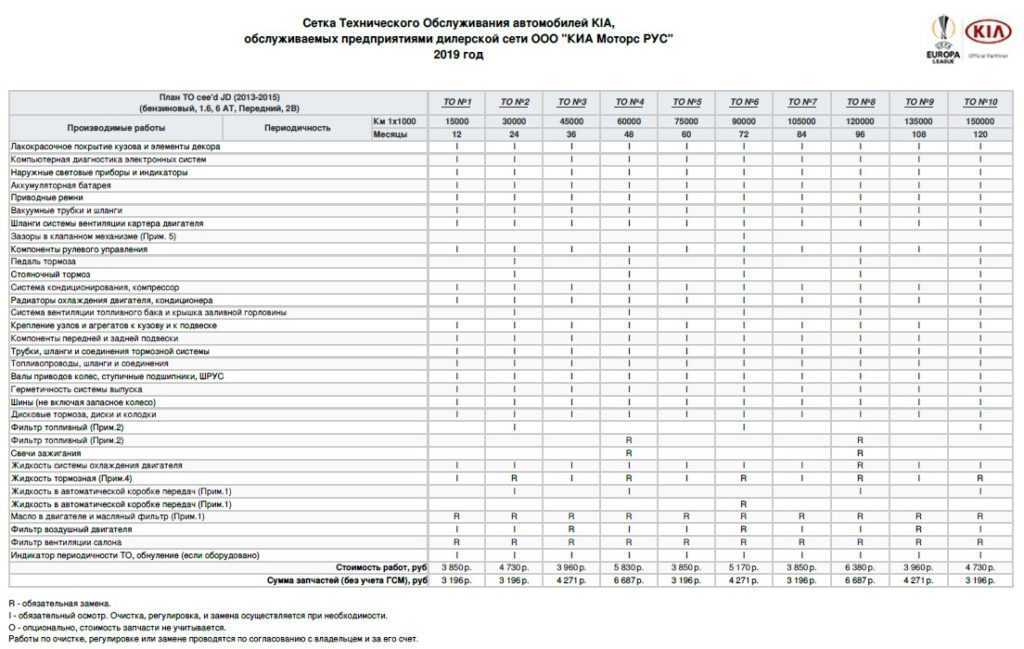
Корреляты выполнения задания
 7). У сытых участников успех диеты был связан с большим количеством ошибок пропуска пищи по сравнению с нейтральными блоками (β = 0,49)., p < 0,001), что не было обнаружено у голодающих участников (β = 0,22, нс ).
7). У сытых участников успех диеты был связан с большим количеством ошибок пропуска пищи по сравнению с нейтральными блоками (β = 0,49)., p < 0,001), что не было обнаружено у голодающих участников (β = 0,22, нс ). Хотя при прогнозировании времени реакции также наблюдалось значительное взаимодействие, бета-веса в обеих группах не были значимыми (βs < 0,24, нс ).
Хотя при прогнозировании времени реакции также наблюдалось значительное взаимодействие, бета-веса в обеих группах не были значимыми (βs < 0,24, нс ). Заключение исследования 2
Текущая тяга к еде
Корреляты текущей тяги к еде
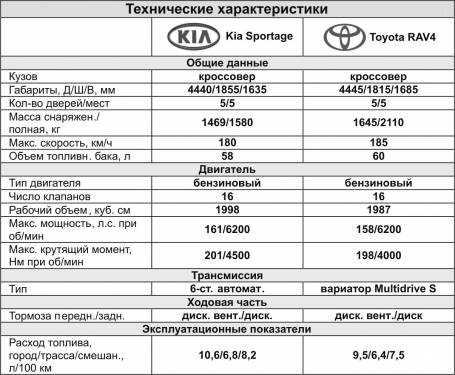 75). В частности, было обнаружено, что довольно неудачные люди, сидящие на диете, увеличивают потребление пищи после потребления высококалорийной предварительной нагрузки (Stroebe, 2008), механизмы которой могут быть аналогичным образом отражены в более высокой тяге, вызванной сигналом к еде, с уменьшением успеха диеты в текущем исследовании. .
75). В частности, было обнаружено, что довольно неудачные люди, сидящие на диете, увеличивают потребление пищи после потребления высококалорийной предварительной нагрузки (Stroebe, 2008), механизмы которой могут быть аналогичным образом отражены в более высокой тяге, вызванной сигналом к еде, с уменьшением успеха диеты в текущем исследовании. . Выполнение задания

Корреляты выполнения задания
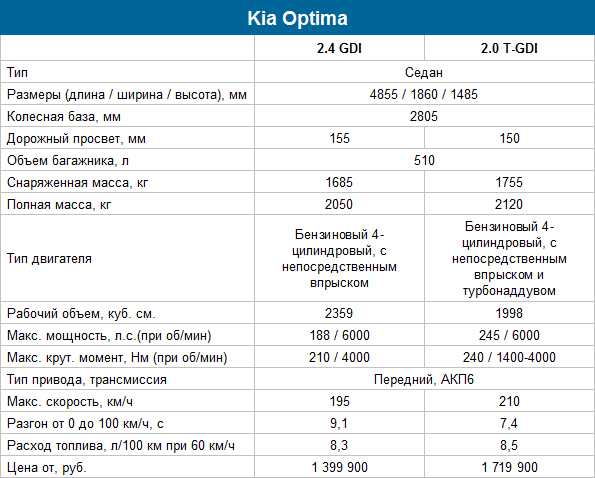 Однако чувство голода и сытости действительно влияет на взаимосвязь между выполнением задачи, успехом диеты и импульсивностью, о которой они сообщают сами себе. В частности, люди с более высоким успехом в диете превзошли тех, кто с меньшим успехом придерживался диеты в пищевых блоках, поскольку они реагировали быстрее, пропускали меньше целей и совершали меньше ошибок, но только когда голодны. Более того, более высокая импульсивность, о которой сообщали сами участники, была связана со снижением поведенческого торможения в нейтральных блоках, но только в состоянии насыщения.
Однако чувство голода и сытости действительно влияет на взаимосвязь между выполнением задачи, успехом диеты и импульсивностью, о которой они сообщают сами себе. В частности, люди с более высоким успехом в диете превзошли тех, кто с меньшим успехом придерживался диеты в пищевых блоках, поскольку они реагировали быстрее, пропускали меньше целей и совершали меньше ошибок, но только когда голодны. Более того, более высокая импульсивность, о которой сообщали сами участники, была связана со снижением поведенческого торможения в нейтральных блоках, но только в состоянии насыщения. Обсуждение
 , 2008, 2011; Loeber et al., 2012). Более быстрое время реакции в ответ на пищевые мишени может отражать склонность к приближению к положительным стимулам. Например, более быстрая реакция на счастливые лица по сравнению с грустными лицами может быть обнаружена в сопоставимой эмоциональной задаче «да/нет» (Schulz et al., 2007). В обоих настоящих исследованиях эти показатели выполнения задачи не были связаны с текущей депривацией пищи и состоянием тяги к еде, а в исследовании 2 не зависели от приема пищи. Таким образом, оказывается, что более быстрые реакции и меньшее количество нарушений внимания в ответ на изображения пищи могут быть связаны с тем фактом, что они, как правило, являются значимыми стимулами из-за их мотивационной значимости для выживания и, таким образом, более значимы, чем другие стимулы, даже в отсутствие голод. Напротив, в недавнем исследовании, проведенном Loeber et al., количество ошибок пропусков в пищевых блоках было уменьшено с более высоким самоотчетом о голоде. (2013).
, 2008, 2011; Loeber et al., 2012). Более быстрое время реакции в ответ на пищевые мишени может отражать склонность к приближению к положительным стимулам. Например, более быстрая реакция на счастливые лица по сравнению с грустными лицами может быть обнаружена в сопоставимой эмоциональной задаче «да/нет» (Schulz et al., 2007). В обоих настоящих исследованиях эти показатели выполнения задачи не были связаны с текущей депривацией пищи и состоянием тяги к еде, а в исследовании 2 не зависели от приема пищи. Таким образом, оказывается, что более быстрые реакции и меньшее количество нарушений внимания в ответ на изображения пищи могут быть связаны с тем фактом, что они, как правило, являются значимыми стимулами из-за их мотивационной значимости для выживания и, таким образом, более значимы, чем другие стимулы, даже в отсутствие голод. Напротив, в недавнем исследовании, проведенном Loeber et al., количество ошибок пропусков в пищевых блоках было уменьшено с более высоким самоотчетом о голоде. (2013).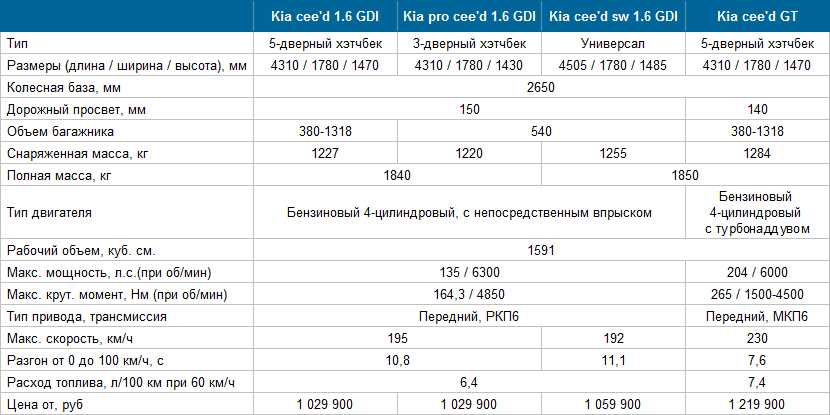 Однако в этом исследовании ошибки пропуска не были связаны с текущими уровнями глюкозы в крови, что предполагает, что — в соответствии с текущими результатами — физиологический механизм из-за дефицита энергии вряд ли может объяснить эти поведенческие реакции.
Однако в этом исследовании ошибки пропуска не были связаны с текущими уровнями глюкозы в крови, что предполагает, что — в соответствии с текущими результатами — физиологический механизм из-за дефицита энергии вряд ли может объяснить эти поведенческие реакции.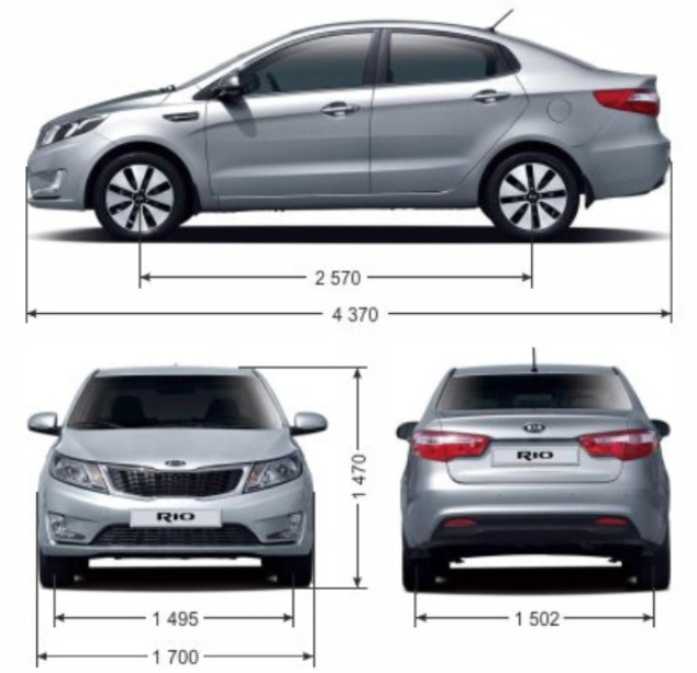 В исследовании 1 комиссионные ошибки не отличались между типами целей. В исследовании 2 участники совершали меньше ошибок в еде, чем в нейтральных блоках. Результаты, вероятно, отличаются, потому что мы увеличили общее количество испытаний в исследовании 2, и впоследствии участники совершили больше ошибок. Повышенное количество комиссионных ошибок в нейтральных блоках (т. е. ложные реакции на пищевые дистракторы) повторяет результаты лексического AST Loeber et al. (2012, 2013). Таким образом, похоже, что пищевые сигналы запускают тенденции автоматических действий, которые могут быть связаны с ярко выраженным стимулом. На ошибки комиссии не влияли пищевая депривация, состояние тяги к еде или потребление пищи в настоящих исследованиях, но они были связаны с самооценкой чувства голода и не зависели от уровня глюкозы в крови в исследовании Loeber et al. (2013). Таким образом, оказывается, что торможение реакции, вызванное пищевым сигналом, аналогично времени реакции и ошибкам пропуска, также не зависит от физиологических процессов из-за текущего статуса питания.
В исследовании 1 комиссионные ошибки не отличались между типами целей. В исследовании 2 участники совершали меньше ошибок в еде, чем в нейтральных блоках. Результаты, вероятно, отличаются, потому что мы увеличили общее количество испытаний в исследовании 2, и впоследствии участники совершили больше ошибок. Повышенное количество комиссионных ошибок в нейтральных блоках (т. е. ложные реакции на пищевые дистракторы) повторяет результаты лексического AST Loeber et al. (2012, 2013). Таким образом, похоже, что пищевые сигналы запускают тенденции автоматических действий, которые могут быть связаны с ярко выраженным стимулом. На ошибки комиссии не влияли пищевая депривация, состояние тяги к еде или потребление пищи в настоящих исследованиях, но они были связаны с самооценкой чувства голода и не зависели от уровня глюкозы в крови в исследовании Loeber et al. (2013). Таким образом, оказывается, что торможение реакции, вызванное пищевым сигналом, аналогично времени реакции и ошибкам пропуска, также не зависит от физиологических процессов из-за текущего статуса питания. Однако это может зависеть от субъективно воспринимаемого чувства голода (по крайней мере, когда оно оценивается Великие весы голода , ср. Лебер и др., 2013).
Однако это может зависеть от субъективно воспринимаемого чувства голода (по крайней мере, когда оно оценивается Великие весы голода , ср. Лебер и др., 2013).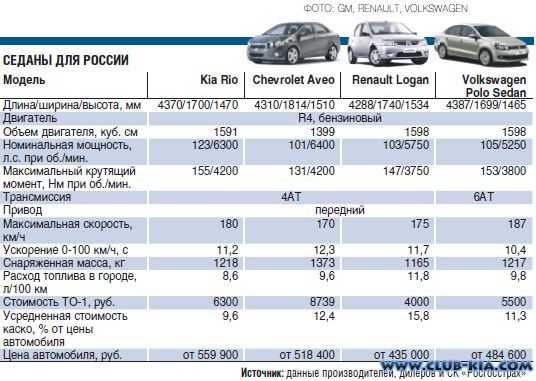 , 2013; Van Koningsbruggen et al., 2013), что может отражаться в повышенном ингибирующем контроле пищевых блоков в текущих исследованиях.
, 2013; Van Koningsbruggen et al., 2013), что может отражаться в повышенном ингибирующем контроле пищевых блоков в текущих исследованиях.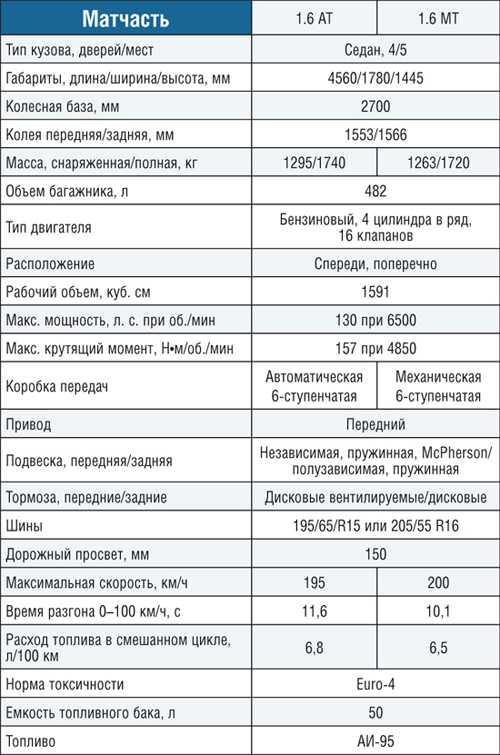 В заключение, хотя связь между торможением реакции, связанной с едой, и успехом диеты не является однозначной, результаты показывают, что выполнение задачи не является независимым от успеха диеты и что отношения могут быть изменены голодом и насыщением.
В заключение, хотя связь между торможением реакции, связанной с едой, и успехом диеты не является однозначной, результаты показывают, что выполнение задачи не является независимым от успеха диеты и что отношения могут быть изменены голодом и насыщением. Текущие результаты показывают, что текущий статус питания может быть важным модератором взаимосвязи между чертой импульсивности и поведенческими реакциями на сигналы, связанные с веществами. В частности, в то время как в большинстве исследований участников обычно инструктируют об умеренном голодании, при исследовании сытых участников могут быть обнаружены более тесные связи между чертой импульсивности и показателями, связанными с едой, такими как предвзятость внимания, чем сообщалось ранее. Это может быть частично объяснено тем фактом, что влияние ситуационных факторов, таких как механизмы гомеостатического голода, снижается, тем самым увеличивая влияние показателей, связанных с чертами характера, таких как самооценка импульсивности (Lowe and Butryn, 2007; Lowe, 2009).; Коскунпинар и Сайдерс, 2013). В результате предвзятость внимания, связанная с едой, или поведенческая расторможенность, вызванная пищевыми сигналами, может быть выявлена как важный посредник между чертой импульсивности и перееданием (Hou et al.
Текущие результаты показывают, что текущий статус питания может быть важным модератором взаимосвязи между чертой импульсивности и поведенческими реакциями на сигналы, связанные с веществами. В частности, в то время как в большинстве исследований участников обычно инструктируют об умеренном голодании, при исследовании сытых участников могут быть обнаружены более тесные связи между чертой импульсивности и показателями, связанными с едой, такими как предвзятость внимания, чем сообщалось ранее. Это может быть частично объяснено тем фактом, что влияние ситуационных факторов, таких как механизмы гомеостатического голода, снижается, тем самым увеличивая влияние показателей, связанных с чертами характера, таких как самооценка импульсивности (Lowe and Butryn, 2007; Lowe, 2009).; Коскунпинар и Сайдерс, 2013). В результате предвзятость внимания, связанная с едой, или поведенческая расторможенность, вызванная пищевыми сигналами, может быть выявлена как важный посредник между чертой импульсивности и перееданием (Hou et al. , 2011; Meule, 2013).
, 2011; Meule, 2013). Future Directions
 Nederkoorn et al., 2012) или контрольное условие, связанное с пищевыми продуктами, например, низкокалорийные продукты питания (см. Meule и Кюблер, в редакции). В результате нельзя исключить влияние возможного эффекта размера категории или подобного, и необходимы будущие исследования, в которых это будет учтено.
Nederkoorn et al., 2012) или контрольное условие, связанное с пищевыми продуктами, например, низкокалорийные продукты питания (см. Meule и Кюблер, в редакции). В результате нельзя исключить влияние возможного эффекта размера категории или подобного, и необходимы будущие исследования, в которых это будет учтено. , 2011) и влияет на выбор продуктов, так что участники чаще выбирают здоровые закуски и меньше сладостей ( Veling et al., 2013a; Van Koningsbruggen et al., в печати). Предполагается, что механизм таких вмешательств заключается в снижении имплицитных аффективных реакций на соблазнительные стимулы, а не в усилении торможения реакции (Houben et al., 2012a; Veling et al., 2013b). К сожалению, ни одно из этих исследований не оценивало, действительно ли те люди, чье потребление пищи было изменено после тренировки, действительно демонстрировали поведенческую расторможенность в ответ на соответствующие пищевые стимулы. Таким образом, в будущих исследованиях может быть изучено, снижает ли такое обучение предвзятость к еде, оцениваемую с помощью AST, то есть усиливает поведенческое торможение в пищевых блоках, и может ли это изменение поведения также привести к долгосрочному снижению потребления нездоровой пищи, а также к более серьезным последствиям. выбор здоровой пищи, особенно у неудачников, сидящих на диете.
, 2011) и влияет на выбор продуктов, так что участники чаще выбирают здоровые закуски и меньше сладостей ( Veling et al., 2013a; Van Koningsbruggen et al., в печати). Предполагается, что механизм таких вмешательств заключается в снижении имплицитных аффективных реакций на соблазнительные стимулы, а не в усилении торможения реакции (Houben et al., 2012a; Veling et al., 2013b). К сожалению, ни одно из этих исследований не оценивало, действительно ли те люди, чье потребление пищи было изменено после тренировки, действительно демонстрировали поведенческую расторможенность в ответ на соответствующие пищевые стимулы. Таким образом, в будущих исследованиях может быть изучено, снижает ли такое обучение предвзятость к еде, оцениваемую с помощью AST, то есть усиливает поведенческое торможение в пищевых блоках, и может ли это изменение поведения также привести к долгосрочному снижению потребления нездоровой пищи, а также к более серьезным последствиям. выбор здоровой пищи, особенно у неудачников, сидящих на диете.
Заявление о конфликте интересов
Благодарности
 DFG не участвовал в разработке исследования, сборе, анализе или интерпретации данных, написании рукописи или принятии решения о подаче статьи для публикации. 9Сообщенные данные были частью исследования, которое также включало другие задачи и физиологические записи в трех случаях, о которых сообщается в другом месте (Meule et al., 2012b, d, 2014). Порядок заданий был уравновешенным, т. е. испытуемые выполняли ТЧД либо на первом, либо на втором, либо на третьем занятии. Каждый сеанс проводился в отдельные дни с интервалом в 1 неделю, и ни один из стимулов, используемых в AST, не использовался в других задачах. Таким образом, мы утверждаем, что маловероятно, что выполнение других задач отрицательно повлияло на результаты, полученные с помощью ТЧА. 9Обратите внимание, что результаты для каждой подшкалы FCQ-S (т.е. желание, подкрепление, облегчение, отсутствие контроля, голод ) были аналогичны результатам для общего балла. То есть баллы по каждой подшкале снижались после приема пищи в сытой группе, были ниже до и после задания в сытой группе по сравнению с голодной и увеличивались в обеих группах после задания по сравнению с до (всего пс).
DFG не участвовал в разработке исследования, сборе, анализе или интерпретации данных, написании рукописи или принятии решения о подаче статьи для публикации. 9Сообщенные данные были частью исследования, которое также включало другие задачи и физиологические записи в трех случаях, о которых сообщается в другом месте (Meule et al., 2012b, d, 2014). Порядок заданий был уравновешенным, т. е. испытуемые выполняли ТЧД либо на первом, либо на втором, либо на третьем занятии. Каждый сеанс проводился в отдельные дни с интервалом в 1 неделю, и ни один из стимулов, используемых в AST, не использовался в других задачах. Таким образом, мы утверждаем, что маловероятно, что выполнение других задач отрицательно повлияло на результаты, полученные с помощью ТЧА. 9Обратите внимание, что результаты для каждой подшкалы FCQ-S (т.е. желание, подкрепление, облегчение, отсутствие контроля, голод ) были аналогичны результатам для общего балла. То есть баллы по каждой подшкале снижались после приема пищи в сытой группе, были ниже до и после задания в сытой группе по сравнению с голодной и увеличивались в обеих группах после задания по сравнению с до (всего пс).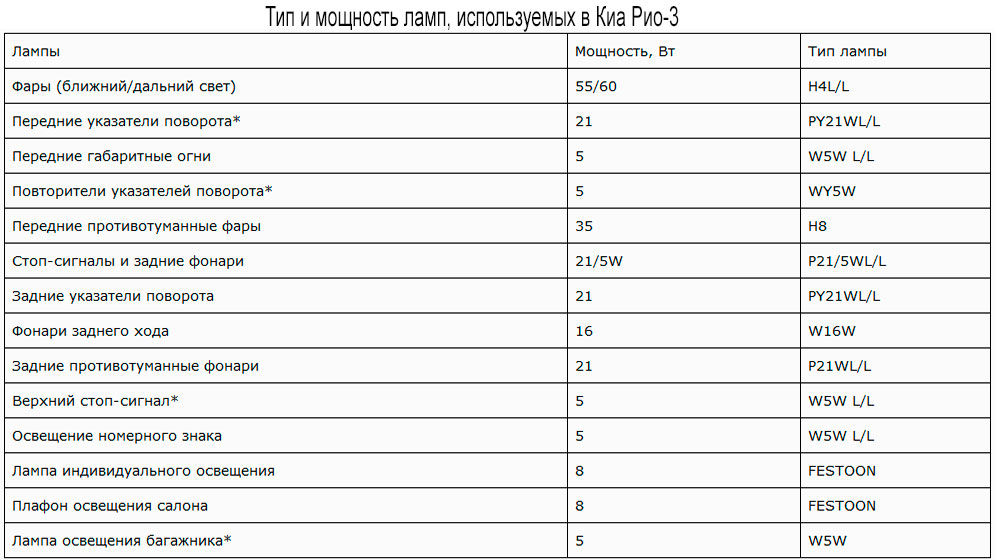 < 0,01).
< 0,01). Ссылки
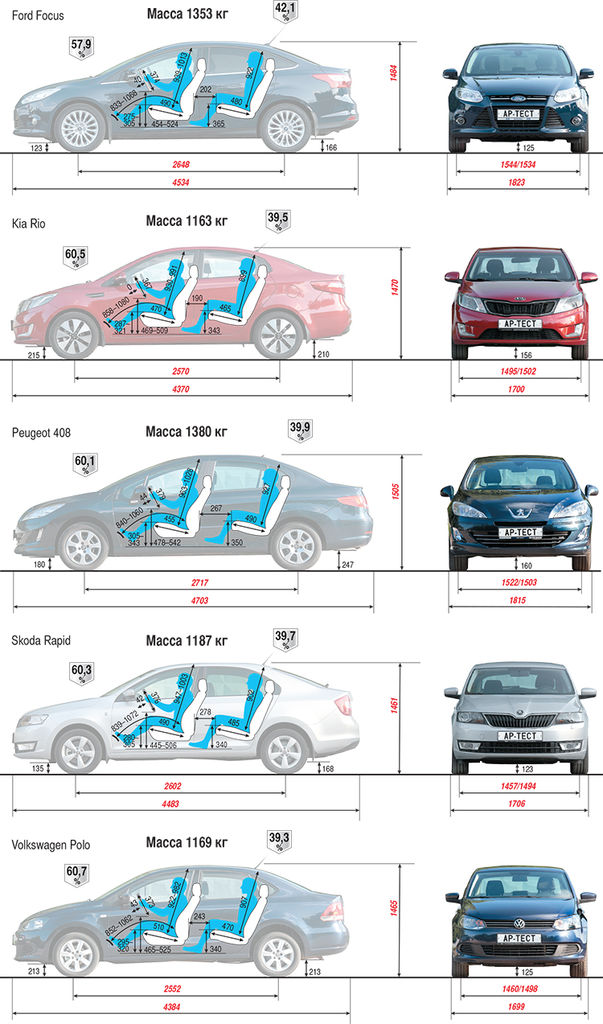 М., Вульф, К., Пагото, С.Л., Шнайдер, К.Л., Уайтд, М.С., и Либман, Р. (2011). Ингибирование пищевого вознаграждения: задержка дисконтирования, чувствительность к пищевому вознаграждению и прием вкусной пищи у женщин с избыточным весом и ожирением. Ожирение 19, 2175–2182. doi: 10.1038/oby.2011.57
М., Вульф, К., Пагото, С.Л., Шнайдер, К.Л., Уайтд, М.С., и Либман, Р. (2011). Ингибирование пищевого вознаграждения: задержка дисконтирования, чувствительность к пищевому вознаграждению и прием вкусной пищи у женщин с избыточным весом и ожирением. Ожирение 19, 2175–2182. doi: 10.1038/oby.2011.57 , Гибсон С., Бенсон Л., Охнер С. Н. и Гелибтер А. (2012). Нейровизуализация и ожирение: современные знания и направления на будущее. Обес. Версия . 13, 43–56. doi: 10.1111/j.1467-789X.2011.00927.x
, Гибсон С., Бенсон Л., Охнер С. Н. и Гелибтер А. (2012). Нейровизуализация и ожирение: современные знания и направления на будущее. Обес. Версия . 13, 43–56. doi: 10.1111/j.1467-789X.2011.00927.x Междунар. Дж. Ешьте. Беспорядок . 45, 407–414. doi: 10.1002/eat.20966
Междунар. Дж. Ешьте. Беспорядок . 45, 407–414. doi: 10.1002/eat.20966 С., Гонсалес-Хименес, Э., Кампой, К., и Вердехо-Гарсия, А. (2012). ИМТ предсказывает эмоциональную импульсивность и когнитивную негибкость у подростков с избыточным весом. Ожирение 20, 1604–1610. doi: 10.1038/oby.2012.47
С., Гонсалес-Хименес, Э., Кампой, К., и Вердехо-Гарсия, А. (2012). ИМТ предсказывает эмоциональную импульсивность и когнитивную негибкость у подростков с избыточным весом. Ожирение 20, 1604–1610. doi: 10.1038/oby.2012.47 84, 296–309. doi: 10.1037/0022-3514.84.2.296
84, 296–309. doi: 10.1037/0022-3514.84.2.296 , Мартейн К. и Янсен А. (2009). Побуждение к импульсивности приводит к перееданию людей с высоким и низким уровнем сдержанности в еде, в то время как те, кто сидит на диете, придерживаются своей диеты. Аппетит 53, 93–100. doi: 10.1016/j.appet.2009.05.013
, Мартейн К. и Янсен А. (2009). Побуждение к импульсивности приводит к перееданию людей с высоким и низким уровнем сдержанности в еде, в то время как те, кто сидит на диете, придерживаются своей диеты. Аппетит 53, 93–100. doi: 10.1016/j.appet.2009.05.013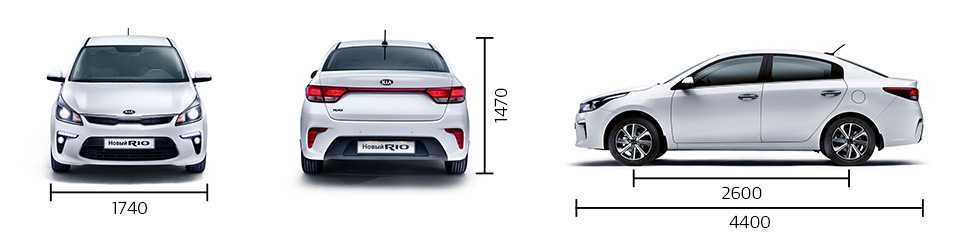 П., и Мак, Д. (1975). Сдержанное и безудержное питание. Дж. Перс . 43, 647–660. doi: 10.1111/j.1467-6494.1975.tb00727.x
П., и Мак, Д. (1975). Сдержанное и безудержное питание. Дж. Перс . 43, 647–660. doi: 10.1111/j.1467-6494.1975.tb00727.x 01.019
01.019 , Nederkoorn, C., and Jansen, A. (2012b). Слишком заманчиво, чтобы сопротивляться? Прошлые успехи в контроле веса, а не диетические ограничения, определяют расторможенное питание, вызванное воздействием. Аппетит 59, 550–555. doi: 10.1016/j.appet.2012.07.004
, Nederkoorn, C., and Jansen, A. (2012b). Слишком заманчиво, чтобы сопротивляться? Прошлые успехи в контроле веса, а не диетические ограничения, определяют расторможенное питание, вызванное воздействием. Аппетит 59, 550–555. doi: 10.1016/j.appet.2012.07.004 Дж., Ясуда, М., Бурант, К. Ф., Грегор, Н., Хатри, С., Свит, М., и др. (2012). Импульсивность и дефицит сдерживающего контроля связаны с нездоровым питанием у молодых людей. Аппетит 59, 738–747. doi: 10.1016/j.appet.2012.08.001
Дж., Ясуда, М., Бурант, К. Ф., Грегор, Н., Хатри, С., Свит, М., и др. (2012). Импульсивность и дефицит сдерживающего контроля связаны с нездоровым питанием у молодых людей. Аппетит 59, 738–747. doi: 10.1016/j.appet.2012.08.001 Аппетит 71, 32–39. doi: 10.1016/j.appet.2013.07.008
Аппетит 71, 32–39. doi: 10.1016/j.appet.2013.07.008 Р. (2009). Комментарий к: «Нейроповеденческое торможение кормления, основанного на вознаграждении: последствия для диеты и ожирения». Ожирение 17, 622–624. doi: 10.1038/oby.2008.639
Р. (2009). Комментарий к: «Нейроповеденческое торможение кормления, основанного на вознаграждении: последствия для диеты и ожирения». Ожирение 17, 622–624. doi: 10.1038/oby.2008.639 , Vögele, C., and Kübler, A. (2011a). Усиленное поведенческое торможение у сдержанных едоков. Ешьте. Поведение . 12, 152–155. doi: 10.1016/j.eatbeh.2011.01.006
, Vögele, C., and Kübler, A. (2011a). Усиленное поведенческое торможение у сдержанных едоков. Ешьте. Поведение . 12, 152–155. doi: 10.1016/j.eatbeh.2011.01.006 Ешьте. Поведение . 13, 423–428. doi: 10.1016/j.eatbeh.2012.08.001
Ешьте. Поведение . 13, 423–428. doi: 10.1016/j.eatbeh.2012.08.001 doi: 10.1016/j.appet.2012.05.010 , Ван дер Линден, М., д’Акремон, М., и Перро, А. (2008). Когнитивный дефицит и предубеждения в отношении еды и тела при булимии: исследование с использованием задания на аффективное переключение. Ешьте. Поведение . 9, 455–461. doi: 10.1016/j.eatbeh.2008.07.002 , Coelho, J.S., Guerrieri, R., Houben, K., and Jansen, A. (2012). Специфика неспособности тормозить реакции у детей с избыточной массой тела. Аппетит 59, 409–413. doi: 10.1016/j.appet.2012.05.028 T.Y., Havermans, R., Roefs, A., and Jansen, A. (2006b). Импульсивность у полных женщин. Аппетит 47, 253–256. doi: 10.1016/j.appet.2006.05.008 , Van der Linden, M., d’Acremont, M., Colmant, M., Hanak, C., Pelc, I., et al. (2005). Когнитивные предубеждения в отношении слов, связанных с алкоголем, и дефицит исполнительных функций у полинаркоманов с алкоголизмом. Дополнение 100, 1302–1309. doi: 10.1111/j.1360-0443.2005.01125.x Психология. Бык . 99, 247–262. doi: 10.1037/0033-2909.99.2.247 doi: 10.1037/11753-005 Дж. Клин. Эксп. Нейропсихология . 34, 67–77. doi: 10.1080/13803395.2011.621891 Бр. J. Health Psychol . doi: 10.1111/bjhp.12075 , Engel, D., Holst, C., Lorenz, J., Peacock, M., Stubbs, J., et al. (2013). Когнитивные и связанные с весом корреляты гибкого и жесткого сдержанного пищевого поведения. Ешьте. Поведение . 14, 69–72. doi: 10.1016/j.eatbeh.2012.10.015
doi: 10.1016/j.appet.2012.05.010 , Ван дер Линден, М., д’Акремон, М., и Перро, А. (2008). Когнитивный дефицит и предубеждения в отношении еды и тела при булимии: исследование с использованием задания на аффективное переключение. Ешьте. Поведение . 9, 455–461. doi: 10.1016/j.eatbeh.2008.07.002 , Coelho, J.S., Guerrieri, R., Houben, K., and Jansen, A. (2012). Специфика неспособности тормозить реакции у детей с избыточной массой тела. Аппетит 59, 409–413. doi: 10.1016/j.appet.2012.05.028 T.Y., Havermans, R., Roefs, A., and Jansen, A. (2006b). Импульсивность у полных женщин. Аппетит 47, 253–256. doi: 10.1016/j.appet.2006.05.008 , Van der Linden, M., d’Acremont, M., Colmant, M., Hanak, C., Pelc, I., et al. (2005). Когнитивные предубеждения в отношении слов, связанных с алкоголем, и дефицит исполнительных функций у полинаркоманов с алкоголизмом. Дополнение 100, 1302–1309. doi: 10.1111/j.1360-0443.2005.01125.x Психология. Бык . 99, 247–262. doi: 10.1037/0033-2909.99.2.247 doi: 10.1037/11753-005 Дж. Клин. Эксп. Нейропсихология . 34, 67–77. doi: 10.1080/13803395.2011.621891 Бр. J. Health Psychol . doi: 10.1111/bjhp.12075 , Engel, D., Holst, C., Lorenz, J., Peacock, M., Stubbs, J., et al. (2013). Когнитивные и связанные с весом корреляты гибкого и жесткого сдержанного пищевого поведения. Ешьте. Поведение . 14, 69–72. doi: 10.1016/j.eatbeh.2012.10.015 Gilmer Wood Company
00 После тонкой полировки и отделки Satinwood имеет гладкую, шелковистую поверхность с блестящей глубиной. Его также называют Красносердым. В свежесрезанном виде он ярко-красного цвета, который темнеет при выдержке. Очень устойчивая древесина. Высушенный на воздухе. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. Некоторые из этих деревьев вырастают до огромных размеров. Одно из первых бревен, которые мы получили отсюда, имело 3 1/2 фута в диаметре, 18 футов в длину и весило 7 тонн. Он мелкозернистый и имеет гладкую поверхность. Большинство этих изделий имеют вид «мраморного пирога» с выдающимся конечным зерном. Твердый и тяжелый, способный хорошо полироваться. Редкость на американском рынке. Частично высушенный на воздухе. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. 00 00 Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. 00 Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. 00 Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. Он имеет красивый цвет и зернистость, сладкий запах, очень прост в обработке и стабилен в использовании. Он очень близок к настоящему рио или бразильскому палисандру (Dalbergia nigra) и поэтому очень востребован для музыкальных инструментов, особенно для гитар. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. Это эбеновое дерево имеет заметную устойчивость к растрескиванию, что характерно для некоторых индийских и азиатских видов. Он имеет прекрасную текстуру и легко поддается резке, строганию, точению и фрезерованию. Он довольно мелкозернистый и хорошо полируется. Это черное дерево обычно происходит из Камеруна или Нигерии, а бревна грубо срублены в лесу и часто доставляются до ближайшей дороги людьми-носильщиками. С исчезновением других видов черного черного дерева этот вид быстро становится предпочтительным черным деревом. Блестящий с привлекательным внутренним переливом. Очень твердый, тяжелый, мелкозернистый, практически без пор. Слегка маслянистый на ощупь. Заболонь имеет цвет от соломенного до ярко-желтого и может достигать 2 дюймов в ширину. Африканское черное дерево может весить до 85 фунтов на кубический фут. Оно используется для маркетри, инкрустации, рукояток ножей, игровых призывов, рукояток пистолетов, кия для бильярда. приклады, шахматные фигуры, ювелирные изделия и другие драгоценные предметы декоративно-прикладного искусства. Но в основном его используют для токарной обработки, и он претендует на звание лучшего дерева для токарных работ, хорошо известного как эталон, по которому оцениваются другие виды дерева для токарных работ. токарных станков, характер древесины позволяет очень точно обрабатывать острые резцы, оставляя красивую полированную поверхность. Его стабильность, маслянистая природа, устойчивость к влаге и прекрасные отделочные качества делают его превосходным по сравнению с другими видами древесины.